

 영어
영어 프랑스
프랑스 스페인
스페인 러시아
러시아 한국
한국 일본
일본Ginsenoside의 합성방법에 관한 연구
Ginsenosides are 이ma에서active ingredients 의precious 약효 가 있는herbs such as 인삼그리고미국ginseng. Modern pharmacological 연구have shown that ginsenosideshave good pharmacological activities such as anti-tumor,anti-inflammatory, anti-oxidati에그리고inhibition 의apoptosis [1]. Ginsenosides belong 을이triterpenoid class 그리고are glycoside compounds formed 에 의해이connection 의a glycoside precursor 그리고a sugar. According to 이aglycone, ginsenosidescan be divided into three types: one is 이oleanane-type pentacyclic triterpenesapon에서Ro, whose aglycone is oleanolic acid; the other two are ginsenosides의ginsenol type (such as Rb1, Rb2, Rc, Rd, F2, Rg3, Rh2, etc.) 그리고ginsenosides의the triterpene type (e.g. Re, Rg1, Rg2, Rf, Rh1, etc.). Both types belong to the dammarane-type tetracyclictriterpene saponins그리고account 을the majority 의ginsenosides, making them the ma에서active ingredients. So far, more than 60 ginsenosideshave been isolated 그리고their structures determined. Most 의the ginsenosidemonomers have significant pharmacological 효과그리고show good 전망for new drug development. For example, Rg1그리고Rb1 have anti-aging activity, 그리고Rg3그리고Rh2 have anti-tumor activity [2].
의 내용 ginsenosides에서인삼그리고American ginseng는 상대적으로 낮습니다.한정된 천연자원으로는 의약품과 연구 개발에 대한 계속 증가하는 수요를 충족하기 어렵습니다.더구나 장기적으로 의약원료로서 야생자원에 의존하게 되면 필연적으로 자연자원의 고갈을 초래하게 되고 생태균형을 심각하게 손상시키게 된다.인삼과 미국인삼의 인공재배는 4~15년의 생장과 재배주기가 필요하며 농약 잔류량, 중금속 오염, 종육 저하 등 문제에 직면해 있다.이러한것들은 모두 대규모적인 경작을 제약하는것입니다.부족한 약용자원의 딜레마를 해소하고 동시에 소중한 자연자원을 보호하기 위해 최근 국내외 학자들은 조직배양, 생물변형, 합성생물학을 통해 진세노사이드의 생합성에 대해 연구했고, 일부 성과를 거두었다.본 논문에서는 이러한 발전상을 검토하고 ginsenosides의 생합성에 대한 전망을 전망하였다.

인삼 자원 부족 문제 해결을 위한 조직배양을 이용한 연구 1
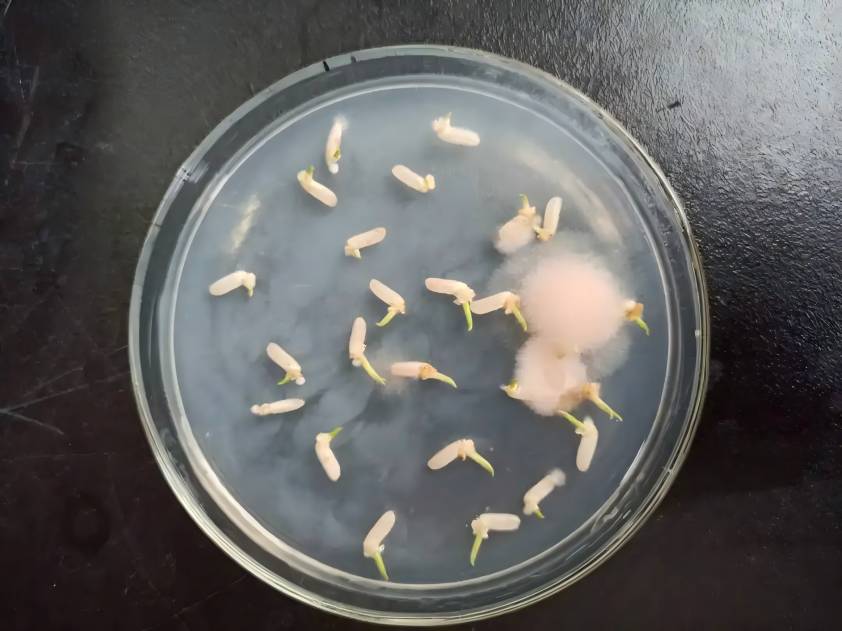
Since the mid-20th century, many researchers have been devoted to the 연구의tissue 문화의인삼그리고American ginseng, attempting to solve the problem 의resource supply of these precious medicinal materials through tissue 그리고셀culture, or to make rapid propagation possible through the obtainment of test tube seedlings. Regarding the 연구of tissue culture of 인삼and American ginseng, there have been many review articles 에서recent years [3, 4]. 식물tissue and 셀culture must ultimately establish large-scale culture methods – reactor culture – 에서order to achieve industrial production. Japan has successfully carried out ginseng cell culture on a 20 000 L bioreactor at a rate of 100 kg per month for the 생산of food additives, and the composition of the ginsenosides 에culture is almost exactly the same as that of cultivated ginseng roots [5]. South Korea has also made great 진행에서the fermentation culture of ginseng adventitious roots and fibrous roots, and the scale of the reactor has reached 10,000 L [6]. 중국has developed the Ding Jiayi series of cosmetics through ginseng callus culture [7].
인삼과 미국인삼의 조직 및 세포배양 연구가 비교적 심도 있게 진행되었고, 세포배양, 섬유근 및 크라운쓸개종양배양, 반응기배양 등에서 어느 정도 진전이 있었지만, 그러나 아직 세포주가 불안정하고, 스케일업 배양이 어려우며, 대상 제품의 함량이 낮고, 복잡한 배양 조건으로 인해 비용이 많이 드는 등의 문제점이 있다.따라서 people&를 충족시키기 위해서는 산업화가 용이한보다 적합한 문화조건과 기술방법을 찾아야 한다#39;s 제약 및 인삼 및 미국 인삼 약재 및 ginsenosides에 대한 연구 및 개발 필요.
생물형질을 통한 희귀 진세노사이드 및 아글리콘 획득 연구 2
Panax 식물에는 다양한 ginsenosides 가 서로 다른 양으로 존재하며 서로 다른 약리학적 기능을 가지고 있다.Rb1, Rg1과 같은 일부 진세노사이드 (ginsenoside)는 매우 풍부한 반면, Rh1, Rh2, Rh3, Rg3와 같은 다른 것들은 매우 소량 존재하는 희귀 진세노사이드 (ginsenoside)이다 [2].많은 약리학적 연구에 따르면 희귀한 진세노사이드가 일반적으로 더 나은 약리학적 활성을 가지고 있다.ginsenosideprotopanaxadiol은 항암활성이 가장 강하며, 당 염기의 수가 증가할수록 ginsenoside의 항암활성이 차례로 감소하여 protopanaxadiol >단당류 글리코사이드>이당류 글리코사이드>삼당류 glycosides >테트라당류 글리코사이드 [8].또한 체내 진세노사이드 (ginsenoside)의 대사 패턴을 연구하여, 대부분의 진세노사이드 (ginsenoside) 가 위장관에서 잘 흡수되지 않으며, 탈당된 2차 진세노사이드 (secondary ginsenoside)와 탈당된 진세노사이드 (ginsenoside) 가 진세노사이드 (ginsenoside)보다 약리학적 활성이 강하고 생체이용성이 높다는 것을 확인하였다 [9-11].
Rare ginsenosides and aglycones are present 에서very small amounts 에서the raw ginseng plant, cell cultures, adventitious roots and fibrous roots. 에서the past, they were mainly obtained 에 의해physically and chemically hydrolyzing the glycosidic bonds of ginsenosides, but the hydrolysis conditions were harsh, not easy to control, and produced many reaction by-products. In addition, a large amount of pollutants such as organic solvents, acids and alkalis were emitted, causing great harm to the environment. In order to avoid damaging the ecological environment and to obta에서resources for sustainable use, many researchers have used bio변환pathways to modify the sugar cha에서structure of ginsenosides 에서order to obtain rare ginsenosides and aglycones [12].
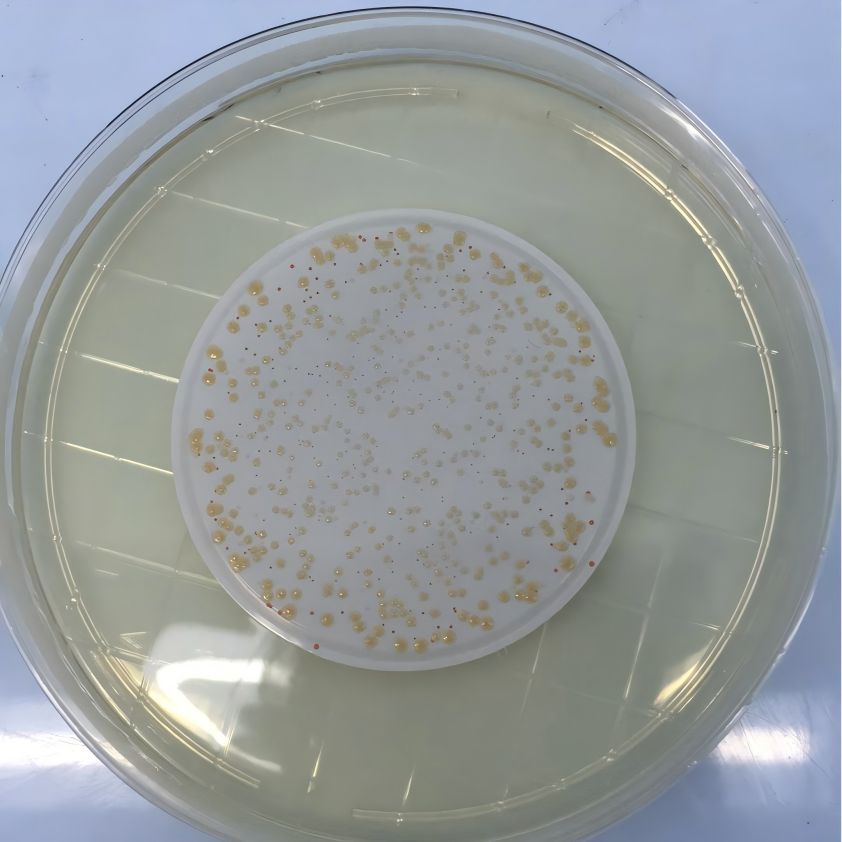
생물전환 (生物轉換, Bioconversion)은 생물체 (세포, 세포기관) 또는 효소를 촉매로 사용하여 화학적 전환을 이루는 과정이다.생물계 (세균, 곰팡이, 식물조직 등을 포함) 가 외인성 기질의 구조를 변형시키는 화학반응이다.그것의 본질은 생물 자체에서 생산된 효소를 이용한 외인성 기질의 촉매 반응이다.생물학적 형질전환은 반응조건이 온화하고, 부산물이 거의 생성되지 않으며, 가공이 용이하고 환경친화적이라는 장점이 있다.최근 진세노사이드의 생물형질전환에 대한 연구가 증가하고 있으며, 의미 있는 결과들이 많이 나타나고 있다.진세노사이드의 생물학적 형질전환은 주로 미생물이나 효소를 이용하여 진세놀형 진세노사이드 (ginsenoside)의 C3 및 C20위치와 트리올형 진세노사이드 (triol-type ginsenoside)의 C6 및 C20위치에서 당질구조를 변형시켜, 함량이 높은 진세노사이드 (ginsenoside)를 가수분해하여 희귀한 진세노사이드 (ginsenoside)와 아글리콘 (aglycones)으로 방향 전환시킬 수 있다 [13].생물형질전환의 주요방법은 미생물형질전환과 효소형질전환이다.
2.1 미생물의 형질전환
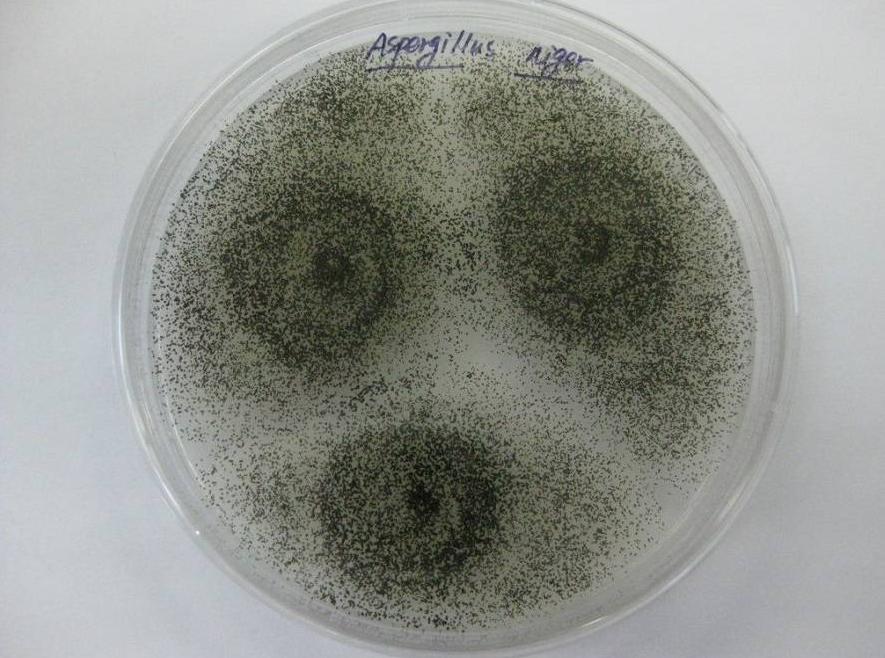
미생물 형질전환 방법은 비용이 저렴하고 부산물이 거의 없으며 널리 사용되고 있다.많은 연구자들이 진세노사이드의 체내 대사를 시뮬레이션한 결과 진세노사이드의 효과를 발휘하는 실제 활성 성분이 일부 장내 식물체를 통해 진세노사이드를 전환함으로써 아글리콘이라는 것을 밝혀냈고, [14]신약 개발의 기초를 마련하였다.Bae 등 15)은 진세노사이드 (ginsenosides)를 컴파운드 K로 전환할 수 있는 유산균을 장내세균으로부터 분리하였으며, 그리고 장내 세균인 Bacteroides sp., Eubacterium sp. 및 Bifidobacterium sp. [16]에 의한 Rg3의 Rh2 및 PPD 로의 전환.진세노사이드를 생체변형시키는 미생물에는 장내 세균 외에도 Aspergillus, Penicillium, Rhizopus, Mucor 종 등의 곰팡이가 있다.동등 [18]은 작은실균류 Aspergillus niger (3.1858)와 Absidia coerulea (3.3538) 가 Rg1을 Rh1로 전환하는 능력이 있으며, 전환율은 80.9%로 나타났다.
푸Jianguo [19]used a large medicinal-food dual-purpose fungus to co-culture with American ginseng and 의root extract, and biotransformed ginsenosides using solid-state fermentation. 이results showed that the enzyme system secreted 에 의해the mycelium has the function of decomposing diol-type saponins, 분해를 가지고 매우 약하 함수를 triterpene saponins, 복합 k. 쳉 외를 생산 할 수 있는 [20]사용 Caulobacter leidyia, β를 생산하는-glucosidase,로 전환하는 ginsenosideRb1 F2 키는 것이다.Bao Haiying et알다.[21]은 Rhizopus sp.를 이용하여 ginsenosideRe를 Rg1, Rg5 및 Rk1로 전환하였으며, 전환율은 최대 92.16%였다.최유등 [22]은 인삼이 재배된 토양에서 4 균주를 분리하였다.인삼열매총사포닌 (SFPG)을 전환하여 컴파운드 K를 전환할 수 있는 Fusarium spp. 균주를 걸러내었다. 이 균주는 높은 특이성과 높은 전환 효율을 가져 컴파운드 K의 산업 생산에 새로운 방법을 제공합니다.
DaiJun-gui et알다.[23, 24] have also done a lot of work on the bio변환of ginsenosides. Rg1and Rb1 were converted 에 의해Fusarium oxysporum Z-001, and a 총of 9 대사 물질were obtained, of which 6α, 12 β-dihydroxydammar-3-one-20 (S) 가 될-β-D-glucopyranoside과 3a-oxa-3a-homo-6 α, 12 β-dihydroxydammar-3-one-20 (S) 가 될-β-D-glucopyranoside들은 새로 운 화합물이다; Rg1 파란 색의 균은 sp에 의해 5 물질로 전환 되었다. AS3.2462, 그중 3-oxo-7 β-hydroxy-20 (S)-protopanaxatriol 그리고 3-oxo-7 β, 15 α-dihydroxy-20 (S)-protopanaxatriol은 새로 운 화합물이다.
2.2 효소 전환
미생물 형질전환과 비교할 때 효소형질전환방법은 반응주기가 짧고 오염이 적으며 얻은 생성물의 순도가 높고 제어성이 강하며 특정성이 있다.서로 다른 성질의 효소들은 서로 다른 형성 및 조성의 글리코시드 결합에 작용하므로 원하는 생성물을 목표로 형성시킬 수 있다.코등 [25]은 다양한 글리코사이드 가수분해효소에 의한 트리테르펜 글리코사이드 혼합물의 가수분해를 연구하였다.…의 혼합물 triterpene saponinsAspergillus oryzae의 galactosidase와 Penicillium sp.의 lactase의 조효소용액을 이용하여 가수분해하였다. 각각, 많은 양의 Rg2와 Rh1을 생산하였다;Penicillium decumbens에서 추출한 naringinase의 조효소추출물을 이용한 트리올형 사포닌 혼합물의 가수분해 결과 장내 대사산물 F1과 소량의 20(S)-PPT 가 생성되었다.본 증례는 triol-type saponin 혼합물의 효소가수분해를 이용한 Rg2, Rh1및 F1의 효율적인 제조에 대한 1차 보고이다.후속 연구의 변환에 diol glycoside 혼합물에 의해 다양 한 glycosidase 효소 원유 효소 솔루션 다는 것을 발견의 락 타 아제에서 균은 oryzae, β에서-galactosidase 및 cellulase Trichoderma viride F2 키 변환 할 수 있 다면, 복합 K및 Rd, 각각; Penicillium 으로부터 lactase의 조제된 효소용액은 Rd, Rg3와 Compound k를 전환할 수 있다. 이는 diol-type saponins의 혼합물을 이용하여 대량으로 F2와 Rg3를 효소적으로 제조한 최초의 보고이다 [26].
류 등 (27)은 Aspergillus niger 로부터 얻어진 crude glycosidase를 이용하여 Rg3 (R)을 PPD (S, R)로 전환하였으며, 전환율은 100%까지;그리고 Rf를 90.4%의 전환율 [28]로 20(S)-PPT로 변환하였다.진펑시 (진Fengxie) [29].연구그룹은 효소전환에 의한 희귀 saponin Rh2 생산에 대해 많은 연구를 수행했습니다.를 사용 하여 새롭게 발견 된 ginsenosideβ-glucosidase,의 glycosyl 그룹 ginsenoside diol 유형이 부분적으로 다른 saponins hydrolyzed Rh2를 생산하고 한다.효소를 생산하는 조작세균을 스크리닝, 배육하여 효소전환산물의 2차 사포닌에 대한 분리 및 정제기술을 개발하였으며, Rh2는 효소적 전환을 통해 산업적으로 생산될 수 있다.이 방법은 ginsenoside diol 로부터 Rh2 생성에 60% 이상의 전환율을 가지는 것으로 생산 실무에서 확인되었으며, Rh2의 순도는 90% 이며, 생삼 원료인 0.5% 이상의 Rh2의 수율을 가지는 것으로 나타나 홍삼으로부터의 수율보다 500배 높은 수율을 나타내었다.반응 후 효소의 회수율은 60%이다.
유전공학 기술의 발달로 최근 일부 연구자들은 글리코시다제 유전자를 대장균으로 전이시키려는 시도를 하기 시작했다.고도발현을 통해 얻어진 재조합 효소에 의한 진세노사이드 (ginsenosides)의 생물학적 형질전환에 어느 정도 진전이 있었다.노 et알다.[30]를 옮 겨 β-glycosidase에서 유전자 복제 Sulfolobus solfataricus 대장균로 변 했다.얻어진 재조합 효소는 전환할 수 있었다인삼 뿌리 추출물화합물 K로, 전환율은 80.5%로.유 et알다.[31]를 옮 겨 β-glycosidase에서 유전자 복제 Pyrococcus furiosus 대장균에, 그리고 그에 따 른 재조 합 효소 첫 화합물에 인삼 추출 루트 변환 된 K의 전환 율을 가 진 79.5%, 그리고 aglycone에 PPD,의 전환 율을 100%,의 수율 ginsenosides에 도달 할 수 있는 1. 8 g · L-1다.본 연구의 Compound K와 ginsenosides의 생산량은 문헌에 가장 높게 보고되어 산업화 가능성이 높다.
진세노사이드 (ginsenosides)의 생물형질전환에는 상당한 진전이 있었지만, 산업화를 이루기 위해서는 특이적으로 형질전환시킬 수 있는 고생산성의 균주 및 효소를 선별하거나 유전공학을 통해 높은 형질전환활성을 갖는 재조합효소를 확보한 후 적합한 산업생산조건을 확립해야 한다.이는 진세노사이드 유도체의 대규모 생산과 혁신적인 약물 연구에 매우 중요한 의미를 지닌다.
진세노사이드에 대한 합성생물학 연구 탐구 3
천연물의약품의 합성생물학은 천연물의약품의 생합성에 관여하는 성분을 발견하여 특성을 규명하고, 이를 공학적 원리를 이용하여 설계 및 표준화하며, 생합성 경로와 대사망을 차대세포에서 조합 및 통합하여 재구성하는 유전체연구,활성 제약 성분의 목표하고 효율적인 이형 합성을 달성하고 천연 의약품 [32]의 연구, 개발 및 제조에서 일련의 주요 문제를 해결하기 위해.천연물 약물 설계 및 합성 분야에서는 합성생물학을 적용하면 대사 경로를보다 정밀하게 제어할 수 있다.천연 제품 생합성 경로의 유전자 조작은 혁신적인 천연 제품 기반 약물 분자를 생산하는 데 사용될 수 있습니다.중요한 천연 약물을 생산할 수 있는 합성"슈퍼 생산자"도 설계 및 시공할 수 있습니다.원하는 목표 화합물은"슈퍼 생산자"를 발효시키는 것만으로 직접 얻을 수 있습니다. 그리고 향후 의약품 생산을 위한 가장 유망한 녹색기술 중 하나가 될 것으로 기대된다.이는 식물로부터 천연약물을 연구, 개발함으로써 초래될수 있는 자원문제를 효과적으로 해결할수 있다.현재 합성생물학은 일부 약용천연물의 제조에서 큰 발전을 가져왔다.
왕Wei 등은 처음으로 중국 주목으로부터 taxane synthase유전자를 클로닝하였고, Saccharomyces cerevisiae에 도입하여 taxane 생합성 경로를 구축하였다.재조합세균은 파클리탁셀의 전구체인 taxane을 직접 생산할 수 있어 taxane 화합물의 합성생물학 연구의 토대를 마련하였다.Ajikumar 등은 Escherichia coli를 이용하여 paclitaxel&의 발효 생산에 성공했다#39;s 키 전구체, taxol.수율은 fed-batch 재배를 통해 1 g·L-1에 도달할 수 있으며, 이는 파클리탁셀 생합성의 다른 단계를 더욱 최적화함으로써 궁극적으로 합성생물학을 통한 파클리탁셀의 대규모 조성이 이루어질 것으로 기대된다.
공장경 등 [35]은 artemisinin 생합성과 관련된 유전자를 Saccharomyces cerevisiae에 전달함으로써 artemisinin의 전구물질인 zhishuihuai-4, 11-diene 및 artemisinic acid를 얻었다.게다가 zhishuihuai-4, 11-diene synthase는 완전히 유전적으로 최적화되어 zhishuihuai-4, 11-diene synthase의 촉매 효율을 현저히 향상시켰으며 조작된 박테리아의 아르테미시닌-4, 11-diene의 수율을 크게 향상시킵니다.Westfall등 36)은 회분식 fed-batch 배양을 통해 40 g·L-1의 수율을 도달할 수 있는 artemisinin-4, 11-diene의 생산을 위한 Saccharomyces cerevisiae 공법 균주를 제조하여 artemisinin의 직접 전구체인 dihydroartemisinic acid로 합성하고, 총 48.4%의 수익률로.이로 인해 합성생물학을 통해 아르테미시닌 전구체를 대량으로 준비할 수 있으며, 아르테미시닌의 합성 경로를 단순화시켜 아르테미시닌의 생산 비용을 크게 절감할 수 있다.
In recent years, research on the biosynthesispathway of ginsenosides and 의reaction mechanism has made some progress, laying the foundation for the 생산of ginsenosides through synthetic biology technology [37, 38].이ginsenoside biosynthesispathway includes more than 20 consecutive enzymatic reactions (Figure 1). The key 효소are 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR), farnesyldiophosphate synthase(FPS), 스쿠알렌synthase(SS), squalene epoxidase (SE), and dammarenediol-II synthase(DDS). FPS), squalene synthase (SS), squalene epoxidase (SE), dammarenediol-II synthase (DS), AS), cytochrome P450(CYP450) and glycosyltransferase (GT), etc.
3. 1 3-Hydroxy-3-methylglutaryl-coenzyme
reductase (HMGR) HMGR은 진세노사이드 생합성 경로에서 최초의 속도 제한 효소로 인식된다.테르페노이드 합성 과정에서 가장 먼저 기능하는 핵심 효소이며, ginsenosides의 전구체인 IPP와 DMAPP의 생성에 영향을 미침으로써 ginsenosides의 생합성에 영향을 미친다.우 씨 등은 4년생 미국 인삼 뿌리에서 HMGR 유전자를 복제했다.암호화된 단백질은 589개의 아미노산으로 이루어져 있다.생물정보학 분석 결과 HMGR은 두 개의 transmembrane domain과 촉매 domain을 포함하고 있는 것으로 나타났다.이 유전자는 많은 식물로부터 복제된 HMGR 유전자와 높은 상동성을 보이며, 특히 동백나무 (Camellia sinensis)의 HMGR 유전자와 상동성이 83.8%까지 나타났다.Camellia sinensis의 monoterpene indole alkaloid인 camptothecin의 생합성은 mevalonate (MVA) 경로를 거쳐야 한다. HMGR 유전자는 ginsenosides의 생합성과 밀접한 관련이 있음을 추론할 수 있다.
3.2 파르네실 피로인산염 합성효소 (FPS)
김등 (40)은 인삼뿌리, PgFPS에서 FPS를 암호화하는 유전자를 복제하였으며, 암호화된 아미노산 서열은 Arabidopsis, rubber, Artemisia annua 및 Centella asiatica의 FPS와 각각 77%, 84%, 87%, 95% 상동성을 보였다.Southern blot 분석 결과 인삼에는 FPS를 암호화하는 유전자가 2개 이상 존재하는 것으로 나타났다.재조합 단백질은 Escherichia coli에서 PgFPS를 발현함으로써 FPS의 활성을 가지는 것을 확인하였다.인삼털뿌리를 methyl jasmonate로 처리한 결과 PgFPS의 mRNA 수준과 FPS의 활성을 모두 증가시키는 것으로 나타났다.이전에 Methyl jasmonate가 인삼 뿌리 정지 세포에서 ginsenoside 축적을 유도하는 것으로 보고되었는데 [41], 이는 PgFPS의 발현 증가와 관련이 있을 가능성이 가장 높다.
김et알다.[42]also overexpressed PgFPS 에서ginseng in hairy roots of Centella asiatica and found that the mRNA level of the damaran synthase in Centella asiatica was significantly increased, and the production of the triterpene saponins madecassoside and asiaticoside increased transiently. The above 연구show that FPS plays an important role in the biosynthesisof triterpenoids and is an important component for improving ginsenoside production using synthetic biology techniques.
3.3 스콸렌 합성효소 (SS)
SS는 아이소프레노이드 경로의 분기점에 위치하며 스테롤과 트리테르페노이드의 합성의 초기 단계를 촉매한다.그 내용과 활동은 진세노사이드 생성에 매우 중요한 역할을 한다.리등 [43]은 인삼 잎으로 제작된 cDNA 라이브러리에서 SS 유전자 PgSS1을 클로닝하였다.이 유전자는 콩, Arabidopsis, 담배 및 쌀에서 각각 84.1%, 75.78%, 81.45% 및 71.33%의 SS 유전자와 상동성을 보이며, 콩, Arabidopsis, 담배 및 쌀에서 각각 84.1%, 75.78%, 81.45% 및 71.33%의 상동성을 보인다.SS 유전자에 대한 식물발현 벡터를 제작하고, 인삼을 변형시켜 우발적인 뿌리를 얻음으로써 유전자의 발현을 상향 조절하였다.그 결과 모든 하류 유전자의 발현이 상향 조절되어 sterols와 ginsenosides의 증가를 가져왔으며, SS 가 sterols와 ginsenosides의 생합성에 조절 역할을 한다는 것을 알 수 있었다.
김등 (44)은 adventitious roots를 재료로 하여 구축한 EST(cardiac sequence tag) 라이브러리에서 두 개의 다른 PgSS1 상동유전자인 PgSS2와 PgSS3를 복제하였다.기능성 보완분석 결과 PgSS1, PgSS2, PgSS3 모두 Saccharomyces cerevisiae SS 유전자 결함 돌연변이의 ergosterol prototrophic 표현형을 복원할 수 있었다.In situ hybridization 분석 결과 인삼의 장기마다 이들 세 유전자의 전사 수준이 다르게 나타났다.이러한 결과는 3개의 SS 유전자가 서로 다른 발현양상을 보이지만 인삼의 스쿠알렌 합성에 모두 관여하고 있음을 나타낸다.서 교수 등 [45]은 인삼의 SS 유전자를 시베리아 인삼의 굳은살 속에 Agrobacterium tumefaciens로 옮겨 최종 대사산물을 발현시켰다.그 결과 인삼 SS의 활성을 강화하면 스테롤의 생성이 유의적으로 증가했으며 트리테르페노이드 사포닌의 생성도 증가하는 것으로 나타났다.따라서 SS는 인삼 사포닌의 합성에 핵심적인 효소임을 추론할 수 있다.SS의 발현을 증가시키면 FPP 가 squalene으로 전환되는 것을 촉진할 뿐만 아니라 다른 하류 효소들의 활성을 상승시켜 스테롤과 트리테르페노이드의 생성을 증가시킨다.
지 앙Shicui 등 (46)은 인삼 SS 유전자를 기반으로 감각 및 반감각 조각 프라이머를 설계하여 SS 유전자 간섭 발현 벡터를 제작하였다.vector는 Agrobacterium-mediated transformation을 통해 인삼 캘러스 조직으로 변형되었다.변형된 캘러스 조직에서의 SS 유전자 발현 수준은 감소되었으며, 사포닌 함량도 변화하였다. 이는 SS 가 ginsenosides의 생합성 경로에서 핵심적인 효소이며, SS 유전자의 발현을 억제하면 ginsenosides의 생성을 조절할 수 있는 것으로 추론된다.따라서 SS는 합성생물학 기술을 이용하여 진세노사이드 (ginsenoside)의 생산을 향상시키는데 매우 중요한 성분이기도하다.
3.4 스쿠알렌 에폭시다제 (SE)
SE는 스테롤과 트리테르페노이드 생합성의 첫 번째 산화적 반응을 촉매하며 그 합성에 있어 속도 제한 효소 중 하나로 간주된다.한강등 [47]은 인삼 잎과 우발 뿌리로 각각 제작된 cDNA 라이브러리에서 두 개의 SE 유전자인 PgSQE1과 PgSQE2를 복제하였다. PgSQE1 RNA interference (RNAi) 기술을 이용하여 형질전환 인삼 뿌리에서의 PgSQE1을 silencing 하면 PgSQE2와 cycloartenol synthase (CAS)의 발현을 유의적으로 상향시켜 스테롤 함량의 증가를 가져올 수 있음을 확인하였다.이러한 결과는 PgSQE1과 PgSQE2는 서로 다른 조절기전을 가지고 있으며, PgSQE1은 ginsenosides의 합성에만 참여하고 sterol 생성에는 관여하지 않음을 나타낸다.지 앙Shicui 등 48)은 미국 인삼의 조직 및 기관에 따른 총 saponin 및 monomer saponin 함량의 차이와 이와 SS 및 SE 유전자의 발현 수준과의 관계를 조사하였다. 그리고 14개의 조직 및 장기의 SS 및 SE 유전자의 발현 정도가 유의한 차이를 보였으며, ginsenosides Re, Rg1, Rb1, Rd및 total ginsenosides의 함량과 유의한 양의 상관관계를 보였다.이는 SS와 SE 가 ginsenoside 합성 경로에서 대단히 중요한 역할을 한다는 것을 보여준다.
3.5 Dammarenediol-II합성효소 (DS)와 a-amyrin 합성효소 (AS)
OSC (oxidosqualene cyclase)에 의해 촉매되는 2,3-oxidosqualene cyclization은 트리테르페노이드 사포닌과 스테롤의 생합성에 있어 핵심 부위이다.OSC는 다유전자군을 형성하고, oxidosqualene cyclization은 골격이 다른 100개 이상의 트리테르페노이드를 생성할 수 있다.OSC 유전자는 다양한 식물로부터 복제되었다.인삼으로부터 ginsenoside 합성과 관련된 OSC 유전자 2개를 클로닝 하였는데, 이는 각각 damarane 형과 oleanane 형의 ginsenoside 합성의 핵심 효소 유전자인 DS와 AS 유전자이다.쿠시로 등 [49]은 인삼털뿌리를 재료로 사용하여 소체를 준비하였고, 체외에서 2, 3-oxo-squalene을 dammarane-II로 순환 시킬 수 있음을 발견했습니다.Tansakul 등 [50]은 OSC 유전자의 보존적 염기서열을 바탕으로 퇴화 프라이머를 설계하고 인삼 뿌리로부터 DS 유전자 PNA를 복제하였다.Saccharomyces cerevisiae로 전달된 후,이 유전자는 dammarane-II의 생산을 촉진시킬 수 있다.
한 등 [51]은 인삼꽃을 이용하여 구축한 EST 라이브러리에서 DS 유전자 DDS를 복제하였으며, 이는 상술한 PNA 유전자 염기서열과 일치한다.DDS 유전자로 변형된 효모는 damarene-II와 hydroxydamarenone; methyl jasmonate는 DDS 유전자의 발현을 상향조정할 수 있다;DDS 유전자를 RNAi 기술로 silencing 하면 인삼 뿌리의 진세노사이드 생성을 원액의 84.5%까지 줄일 수 있습니다.Lee 등 [52]은 DDS 유전자를 담배에 전이시켜 다마라네 형태의 진세노사이드 (ginsenoside)를 생성함으로써 담배모자이크 바이러스에 대한 담배의 저항성을 현저히 향상시켰다.이러한 결과는 DS 가 ginsenosides의 생합성 경로에서 매우 중요한 역할을 하며 합성생물학 기법을 이용하여 dammarane-type ginsenoside를 얻는데 중요한 요소임을 보여준다.2,3-oxidosqualene의 순환 작용을 촉매로 하여 eudesmanolide를 형성하며, 지금까지 oleanane-type ginsenosides의 합성에서 발견된 유일한 핵심 효소이다.
Kushiro등 (53)은 인삼의 털뿌리로부터 AS의 cDNA 서열을 복제하고, PNY를 Saccharomyces cerevisiae로 이동시켜 ginsenoside Rb1의 생성을 촉매하였다.자 오Shoujing 등 [54]도 인삼 뿌리에서 AS 유전자를 클로닝하였고, AS 유전자에 대한 항감각 식물 발현 벡터를 성공적으로 제작하였다.AS 유전자에 대한 antisense발현벡터를 확립하고 antisense RNA 기술을 이용하여 AS 유전자의 발현을 억제함으로써 대사흐름이 주로 다마란 형태의 triterpene saponin 분지쪽으로 향하게 하여 ginsenosides의 함량을 증가시킨다.
3.6시토크롬 P450 (CYP450)
CYP450은 ginsenosides의 triterpene 탄소 골격의 hydroxylation 및 산화와 같은 복잡한 개질을 수행하기 때문에 ginsenoside 생합성 경로의 핵심 효소이다.최근에는 차세대 염기서열분석 기술과 생물정보학 분석을 이용하여, ginsenoside 생합성에 관여하는 CYP450s 관련 유전자들이 가려졌고, 후보 유전자들의 생물학적 기능이 확인되어 [55].ginsenoside 생합성 경로가 추가로 해명되었다.한강등 (56)은 methyl jasmonate-induced 인삼 첨가 뿌리에 대해 transcriptome 염기서열 분석을 수행하였고, 접합, 주석, 증폭 등을 통해 9개의 후보 전장 CYP450 유전자를 얻었다.그 중 CYP716A47 유전자는 그 발현을 upregulating 하여 methyl jasmonate 유도에 반응하였을 뿐만 아니라 SS 유전자를 과발현 시킨 형질전환 인삼식물로 전이된 후 인삼 뿌리에서 ginsenosides의 생성을 증가시켰다.CYP716A47 유전자를 Saccharomyces cerevisiae로 변환시키면, 발현된 재조합 단백질은 다마레네디올-ii의 C-12위치의 하이드록실화를 촉매하여 프로토긴세놀로 전환할 수 있다.DS와 CYP716A47을 동시에 Saccharomyces cerevisiae로 이동시킨 결과, 재조합 균주에서 protopanaxadiol의 생성이 검출되었다.본 보고는 인삼 사포닌 합성에 관여하는 CYP450을 기능적으로 확인한 최초의 보고이다.
첸 Shilin's [57]research group applied high-throughput 454GS FLX sequencing technology to conduct a transcriptome study of ginseng, American ginseng and Panax notoginseng, and mined CYP450 에서a large amount of transcriptome data, providing an important basis for further screening of CYP450 관련 된ginsenoside에서synthesis. 태양et알다.[58].performed high-throughput sequencing on American ginseng roots and 그리고 접합 및 주석을 통해 150 CYP450s를 얻었다.methyl jasmonate 유도 실험을 위하여 발현이 가장 높은 27 CYP450s transcript level을 선발하였다.뿌리, 줄기, 잎, 꽃 조직 중에서 contig00248 transcript 만이 DS와 동일한 발현 패턴을 보였다.contig00248 transcript는 계통학적으로 Arabidopsis CYP88 과에 가깝다.이 글에서는 damascenone-II 또는 protopanaxadiol의 산화를위한 주요 후보 CYP450 으로서 contig00248 transcript를 사용한다.
루오등 [59]은 Panax 노토삼 뿌리에 대하여 고처리량 염기서열을 수행하였으며, 이를 조합하여 15개의 전장 CYP450s를 주석 및 증폭하였다.그 중 Pn00158 transcript는 미국 인삼 후보인 CYP450 contig00248 transcript와 높은 유사성을 가지고 있으며, 기능적으로 확인된 인삼 CYP716A47 아미노산 서열과 97.95%의 높은 상동성을 가지고 있다.Pn00158이 Panax 노토인삼의 ginsenoside 생합성에 관여하는 CYP450일 가능성이 매우 높다고 추론하는 것.한 등 (60)은 메틸자스몬 유발 인삼의 우점근 EST library에서 CYP716A53v2를 클로닝 하였으며, Saccharomyces cerevisiae에서 발현된 재조합 단백질은 protoginsenolide의 C-6 hydroxylation을 촉매하여 protoginsenolide로 전환시킬 수 있다.상기 ginsenoside 관련 CYP450에 대한 연구 진행은 ginsenoside 생합성 경로에 대한 연구를 크게 진전시켰으며 또한 합성생물학 기술을 통한 ginsenoside 생산 탐구에 중요한 성분을 제공하였다.
3.7 글리코실트랜스퍼라제 (GT)
GT에 의해 촉매된 당화 반응은 진세노사이드의 생합성의 마지막 단계이다.주요 과정은 nucleoside diphosphate의 활성 당 분자를 ginsenoside aglycone 기질로 이동하여 글리코사이드 결합을 형성하는 것이다.당화는 진세노사이드 (ginsenoside)의 안정성과 수용성을 증가시킬 수 있으며,이 과정에서 다양성을 결정하기도 한다.GTs는 높은 특이성을 가진 유전자군의 형태로 식물에도 존재한다.다른 설탕 보습제 또는 다른 설탕 수용자에게 전송하기 위해 다른 GTs 가 필요합니다.첸등 [61]은 먼저 인삼털뿌리로부터 GT를 분리하였으며, 분자량이 SDS-PAGE를 이용하여 56.6 kD 임을 확인하였다. 그리고 그 효소적 특성을 선행 연구하였다.ue 등 (62)은 Rd를 Rb-1로 전환할 수 있는 Panax 노토삼 보류세포로부터 a GT를 분리 및 정제하였다.그러나 파낙스 (Panax) 식물에서 GT 유전자를 클로닝한 사례는 보고된 바 없다.Glycosylation은 ginsenoside 생합성 경로에서 가장 하류 단계이며, 심층적인 연구는 가치가 높은 ginsenoside를 선택적으로 얻는데 매우 중요하다.
요약하면, ginsenoside 생합성 경로와 관련 효소의 기본 뼈대 연구에 상당한 진전이 있었습니다.인삼과 미국인삼 및 Panax 속의 다른 식물로부터 ginsenoside 생합성과 관련된 효소를 암호화하는 20개 이상의 유전자가 복제되어 기능적으로 검증되었으며, 합성생물학 기법을 통해 ginsenoside 생산을 위한 기본적인 생물학적 성분을 제공하고 본 연구의 좋은 토대를 마련하였다.
Saccharomyces cerevisiae는 일반적으로 ginsenoside 생합성에 관여하는 주요 효소를 암호화하는 유전자의 기능을 확인하기 위한 섀시 세포로 사용된다.이는 Saccharomyces cerevisiae 가 단순한 영양소를 가진 배양액에서 생장할 수 있고, 생물반응기를 이용한 생산 규모 확대가 용이하며, 선택할 수 있는 영양소가 부족한 여러 종류, 사용할 수 있는 여러 선택 가능한 마커 등 우수한 섀시 세포에 필요한 특성을 가지고 있기 때문이다. 특히 Saccharomyces cerevisiae에 의해 자체 고유의 MVA 경로를 통해 생성되는 2,3-oxidosqualene은 합성 ginsenosides의 전구체이다.이는 Saccharomyces cerevisiae의 ginsenoside 대사 경로 구축에 큰 편의를 제공한다.또한, 일부 ginsenosides의 glycosyl 그룹은 약리학적 효과에 필요하지 않기 때문에, 탈당된 ginsenoside 아글리콘의 약리학적 활성은 심지어 ginsenosides보다 강하다.따라서 합성생물학을 통해 진세노사이드 아글리콘을 직접 얻는 것도 매우 중요하다.

4 결론
Ginseng, American ginseng and their saponins have become a research hotspot due to the growing demand for medicine and research and development. Some progress has been made in these areas. In addition to the artificial cultivation of ginseng and American ginseng, several methods for obtaining ginseng saponins have been reported at home and abroad, including tissue culture, biotrans형성and synthetic biology techniques. Tissue culture is currently an important way to solve the problem of drug sources. Given that the 대사pathway of ginsenosides has gradually become clear, the expression of key enzyme 유전자can be increased through genetic 공학techniques to improve the synthesis of ginsenosides and obtain high-yield cell lines, thereby more effectively alleviating the growing demand for medicine and research and development. Biotransformation has outstanding advantages in obtaining rare ginsenosides and their aglycones, and it may also be possible to obtain ginsenoside derivatives that do not exist in natural plants. Synthetic biology research on ginsenosides is also a promising approach with broad development prospects.
진세노사이드의 생합성은 여러 가지 요인에 의해 조절되는 복잡하고 동적인 과정이다.합성생물학을 통해 진세노사이드를 생산하고자 하는 목적을 달성하기 위해서는 관련 주요 효소들의 복제된 유전자를 적합한 차대세포로 전달하고,이 유전자들을 인위적으로 변형시켜 이형 및 효율적인 발현을 가능하게 할 뿐만 아니라, 전체 대사 네트워크를 가동시키는 스위치를 찾기 위해, 진세노사이드 대사 네트워크를 조절하는 일부 조절 유전자에 대한 심도 있는 마이닝을 수행해야 한다, 이로써 전체 대사경로에서 유전자의 전반적인 발현 수준을 향상시키고 진세노사이드 생산을 더욱 효과적으로 증가시킨다.지금까지 진세노사이드의 합성생물학 연구에서 생물학적 성분을 얻고 그 기능을 검증하는 측면에서 큰 진전이 있었음에도 불구하고, 차대세포의 변형과 대사경로의 조립에 관한 연구는 이제 막 시작되었다고 할 수 있다.따라서 관련 학문들이 힘을 합쳐 공동으로이 연구의 발전을 도모해야 한다.
참조
[1] 그는 DT, 왕 B, 첸 JM이다. 연구 progress 약리 학적에 효과 of ginsenosides [J]다.J 요 녕 대학 Tradit 턱 병원, 2012년, 14:118-121.
[2] 크리스 텐 슨 LP다. Ginsenosides:화학, 생합성, 항문-분해, and 잠재적인 건강 effects [J]다. Adv 음식 Nutr Res,2009, 55:1-99.
[3] Zuo BM, 가 오 WY, Dong YY, et 알다. 진행 of the 약용식물 Panax 인삼의 조직배양 (J. Mod 턱Med, 2012년,14:34-37 쪽.
[4] 류H, Gao WY,Zuo BM 등이 있다. 부제:진행of the tissue culture in Panax quinquefolium L.[J] Mod 턱Med, 2012, 14:1-4 쪽.
[5] 가오 위, 지아 W,듀안 HQ 등이 출연한다. 약용식물 조직배양의 산업화 (J. 중국 JChin Mater Med, 2003, 28:385-390.
[6] 유 KW, 오 WY, 닥터 한 었, et 알다. Jasmonic 산 Panax 인삼 C. A. Meyer [J]의 우발 뿌리 배양에서 ginsenoside 축적을 개선한다. Biochem Eng J,2002, 11:211-215.
[7] 첸W,Gao WY,Jia W, 외. 조직에 대한 연구의 진보와 cell culture in medicinal 식물 파낙 스의 L. [J]다. Chin Tradit Herb Drugs, 2005, 36:616-620 참조.
[8] Dou DQ, Jin L, 첸 YJ다. 발전 and prospects Panax 인삼의 화학성분과 약리학적 활성에 관한 연구 중 (J. J선양광동 제약Univ, 1999, 16:151-156.
[9]. Kobashi K. 장내 세균과 ginsenosides의 약리학적 효과에 대한 관계 (Relation of 장박테리아to pharmacological effects of ginsenosides) Biosci미생물상, 1997, 16:1-7.
[10] 하세가와 H. the의 증명 신비 로운 효능 인삼의:basic and 임상 재판: metabolic 활성화 ginsenoside deglycosylation의 by 장 박테리아 and esterification 지방산 [J]으로. JPharmacol Sci,2004년,95:153-157.
[11] 왕 YZ, 첸 J, 추 SF,et 알다. 개선 in 기억의 쥐 and 증가 of 해마 excitability in 쥐에 의해 ginsenoside Rg1의 metabolites ginsenoside Rh1 그리고 프로토파낙사트리올 [J. J Pharmacol Sci, 2009, 109:504-510.
[12] 류 X, Dai JG다. 발전 in the 의 biotransformation에 대한 연구 ginsenosides [J]다. 인삼 Res, 2010년, 22:19-22.
[13] 장 표시, 첸 XY, Zhao WQ다. 발전 in studies biotransformation에 of ginsenosides [J]다. J Shenyang Pharm Univ, 2008, 25:419-422.
[14] 왕Y,류TH, 왕W, 외. 의 변용에 대한 연구 ginsenoside Rg1 by 장 플 로라 [J]다. 중국 J Chin Mater Med, 2001년,26:188-190.
[15] Bae EA, Choo MK,공원EK 외. 사람에 의한 ginsenoside R(c)의 대사 장내 세균 and 의 관련 항 알레르기 작용 [J. Biol Pharm Bull, 2002, 25:743-747.
[16] Bae EA, 한강MJ, Choo MK 외. 20(S)-및 20(R)-ginsenoside의 신진대사 Rg3 by 인간 intestinal bacteria and 체외 생물 활동과의 관계 [J]. Biol Pharm Bull, 2002, 25:58-63.
[17] 최인 (쿠이YN), 장익 (장YX), 자오 (Zhao YQ). (영어) 발전in studies on prep-aration 희귀 한 ginsenosides biotransformation에 의해 [J]다. Chin Tradit Herb Drugs, 2009, 40:676-680.
[18]. Dong AL,Cui YJ, Guo HZ, 외. 진세노사이드 (ginsenoside)의 미생물학적 변형 Rg1 [J]다. J Chin Pharm Sci, 2001, 10: 114-118.
[19] Fu JG다. 연구 on the 미생물 변환 of [D] Ginsenoside합니다. 창춘:지린농업대학, 2004.
[20] 청 LQ,김 MK,이 JW, et 알다. 변환 주요 ginsenoside Rb1의 진세노사이드 F2로 Caulobacter leidyać-위키낱말사전Biotechnol Lett, 2006년,28:1121-1127.
[21] Bao HY,리L, Zan LF, 외. Ginsenosides Re biotransformation by Rhizopus sp [J]다. Mycosystema, 2010년, 29일:548-554.
[22] Cui Y, 지 앙 BH, 한강 Y, et 알다. 미생물이 변환에 ginsenoside 화합물 K 에서 total saponins in 과일 파낙스 인삼 [J]의. Chin Tradit Herb Drugs, 2007, 38:189-193.
[23] 류 X, 다오 LR, 장 D, et 알다. 미생물이 탈당화 (deglycosylation)와 케톤화 (ketonization) ginsenosides Rg1의 and Rb1은 Fusarium oxysporum [J]에 의한 것이다. J Asian Nat Prod Res, 2011년,13:652-658.
[24] 류 X, 다오 LR, 장 D, et 알다. 미생물이 transformation Absidia coerulea에 의한 ginsenosides-Rg1과 다약물 저항성 종양세포에 대한 대사산물의 반전 활성 (J.피토테라피아, 2011, 82:1313-1317.
[25] 코 시니어, 최KJ), 스즈키 K (스즈키K) 외 효소 준비 ginsenosides Rg2, Rh1, 및 F1 [J]의. Chem Pharm Bull, 2003, 51:404-408.
[26] Ko 시니어, 스즈키 Y, Suzuki K, et 알다. 표시 production ginsenosides Rd, F2, Rg3 및 compound K의 효소제 me-thod [J. Chem Pharm Bull, 2007, 55:1522-1527.
[27] Liu L, Zhu XM,왕 QJ, et 알다. 효소 준비 20 (S, R)-protopanaxadiol by transformation 20 (S, R)-Rg3 에서 검은 ginseng [J]다. Phytochemistry, 2010년, 71: 1514년-1520.
[28] 류 L, 구 LJ, 장들 외. Aspergillus niger에 의한 희귀한 ginsenoside Rf의 20(S)-protopanaxatriol 로의 미생물 전환 (J. 바이오sci Biotechno1 Biochem, 2010년,74:96-100.
[29] Kim DS, 큐 YS,유 HS 외. Ginsenoside 효소 반응으로부터 준비된 Rh2 [J]. J 대련 Inst Light Ind, 2002, 21:112-115.
[30] 노 KH, 아들 JW, Kim HJ, et 알다. Ginsenoside 열안정성 be-ta-glycosidase에 의한 인삼뿌리 추출물로부터 compound K 생산 에서 Sulfolobus solfataricus [J. Biosci Bio-techno1 Biochem, 2009, 73:316-321.
[31] 유 씨는 MH, 염은 SJ, Park CS, et 알다. 생산 의 aglycon protopanaxadiol를 K 화합물을 통해 阝-glycosidase에서 Pyrococcus furiosus) [J. 한 기초 Microbiol Biotechnol, 2011, 89:1019-1028.
[32] 첸 SL, Zhu XX, 리 낭포 성 섬유 증, et 알다. 유전 체 학 and 한의학의 합성생물학 [J. Acta Pharm Sin, 2012, 47:1070-1078.
[33] 왕 W, 멩 C, Zhu P, et 알다. 예비 study 신진대사에 engineering of 이스트 for 생산 taxadiene [J]다.중국Biotechnol, 2005, 25:103-108.
[34] Ajikumar PK, Xiao WH, Tyo KE, 외. Escherichia coli에서 taxol 전구체 과잉생산을 위한 Isoprenoid 경로 최적화 (J. 과학, 2010, 330:70-74.
[35] 콩 JQ, 왕 W, Wang LN, et 알다. The 개선 amorpha-4,11-diene 생산의 yeast-순응하는 변형 [J].JAppl Microb, 2009, 106:941-951.
[36] Westfall PJ, Pitera DJ, 레니 주니어, et 알다. 생산 효모 중의 amorphadiene의, 그리고 항말라리아 작용제 artemisinin의 전구체인 dihydroartemisinic 산로의 전환 [J. Proc Natl Acad Sci USA, 2012, 109:E111-118.
[37] 우 Q, 저 우 YQ, 태양 C, et 알다. Progress in 에 대한 ginsenosides 생합성 및 2차 대사공학의 전망 the production of ginsenosides [J]다. China Biotechnol, 2009, 29:102-108.
[38] 샤 오 밍 "라고, 한강 T, 황 F, et 알다. 발전 in studies ginsenoside에 biosynthesis and its 관련 enzymes [J]다. Chin Tradit Herb Drugs, 2010, 41:1913-1917 참조.
[39] 우 Q, Sun C, Chen SL다. 식별 and 3-hydroxy-3-methylglutaryl coenzyme a reductase 유전자의 발현 분석 (expression 분석of a 3-hydroxy-3-methylglutaryl coenzyme a reductase gene from American ginseng [J]다. 식물 Omics J, 2012, 5:414-420다.
[40] Kim OT,Bang KH, Jung SJ 외. 인삼 farnesyl diphosphatesynthase 유전자의 분자특성 및 methyl jasmonate [J]에 의한 up-reg-ulation. Biol Plant, 2010, 54:47-53.
[41] 알리 MB, 유 KW, 닥터 한 었, et a1.메 틸 jasmonate 그리고 살리 실 acid elicitaion 유도 ginsenosides 축적, 효소 and non-enzymatic 산화 in 서스펜션 문화 Panax 인삼뿌리 in bioreactors [J]다. 식물 셀Rep, 2006, 25:613-620.
[42] Kim OT,Kim SH, Ohyama K, 외. phytosterol의 상향 조절 및 triterpene biosynthesis in Centella asiatica 털 뿌리 overexpressed ginseng farnesyl diphosphate synthase [J]다.식물셀Rep, 2010, 29:403-411.
[43] Lee MH, 정 씨는 문제이, 서태지 JW, et 알다. 향상 된 스쿠알렌을 과발현 시킨 Panax 인삼의 triterpene과 phytosterol 생합성 synthase 유전자 [J]다. 식물 Cell Physiol, 2004, 45:976-984.
[44] Kim TD, Han JY,Huh 외. 표현 및 기능적 특성화 표현and functional characterization 3의 squalene synthase 유전자 과 관련 된 사포 닌 biosynthesis Panax 인삼 [J]에서. Plant 세포생리학, 2011, 52:125-137.
[45] Seo JW, Jeong JH, Shin CG, 외 a1. Eleutherococcus senticosus에서 squa-lene synthase의 과잉 발현은 phytos-terol과 triterpene 축적을 증가시킨다 [J]다. Phytochemistry, 2005, 66:869-877.
[46] Jiang SC, Zhang MZ, Wang Y, et al. 간섭 vector 건설of Panax 인삼의 SQS 유전자와 transformation callus [J]다.J 지린 Univisity, 2011, 49: 1136-1140다.
[47] Han JY, In JG, 권 씨는 YS, et al. 규정 Panax 인삼에서 squalene epoxidase 유전자의 RNA 간섭에 의한 ginsenoside와 phytosterol 생합성에 관한 연구 (J. Phytochemistry, 2010, 71:36-46.
[48] Jiang SC, Liu WC, Wang Y, et al. 상관 의 ginsenoside 축적과 SQS 및 SQE 유전자 발현 사이 다른 장기 Panax quinquefolius [J]의. Chin Tradit Herb Drugs, 2011, 42:579-584.
[49] Kushiro T, 오노 Y, 시부야 M, et a1. In vitro conversion of 2, 3-oxidosqualene into dammarenediol by Panax 인삼 microsomes [J]. Biol Pharm Bull, 1997, 20:292-294.
[50] Tansakul P, 시부야M, Kushiro T, 외 다수 Dammarenediol-II Panax 인삼에서 최초로 ginsenoside 생합성을 위한 전용 효소인 synthase.FEBS Lett, 2006, 580:5143-5149.
[51] Han JY, 권 씨는 YS, 양 DC, et al. Expression and dammarenediol synthase 유전자의 RNA interference 유도 silencing in 파낙스 인삼 [J.식물 Cell Physiol, 2006, 47:1653-1662
[52] Lee MH, Han JY,Kim HJ 외. Dammarenediol-II 생산은 Panax 인삼 Dammarenediol-II를 발현하는 형질전환 담배에 TMV 허용성을 부여한다 synthase [J]다. Plant cell Physiol, 2011, 53:173-182.
[53] Kushiro T, Shibuya M, Ebizuka Y. 阝-Amyrin 고등식물 중에서 가장 인기 있는 triterpene의 형성을 촉매하는 oxidosqualene cyclase의 synthase cloning에 관한 연구 (J. 유로제이생켐, 1998, 256:238-244.
[54] Zhao SJ, Hou CX, Liang YL, 외. 복제의 인삼 阝 자는 유전자 and the construction of its antisense 식물 표현 벡터 [J]다. China Biotechnol, 2008, 28:74-77.
[55] Niu YY, 루오 흠, 황 카레 브. Advances in the study CYP450 중 ginsenosides 생합성에 관여하는 [J. 세계 Sci Technol/Mod Tradit Chin Med Mater Med, 2012, 14:1177-1183.
[56] Han JY, Kim HJ, 권 씨는 YS, et al. The Cyt P450 효소 CYP716A47은 dammarenediol-II 로부터 protopanaxadiol의 형성을 촉매한다 동안 ginsenoside biosynthesis in 파낙스 인삼 [J. 식물세포생리학, 2011, 52:2062-2073.
[57] Chen SL, Luo 흠, Li Y, et 알다. 454 EST analysis Panax 인삼에서 ginsenoside 생합성에 put적으로 관여하는 유전자를 검출한다 [J]. Plant Cell Rep, 2011, 30:1593-1601.
[58] Sun C, Li Y, Wu Q 외. 드 새로 GS FLX 티타늄을 이용한 미국 인삼 뿌리 전사체의 염기서열 분석 (sequencing and analysis of the American 인삼 뿌리 전사체 플랫폼 to 발견 추정 되는 genes involved in ginsenoside biosynthesis [J]다.BMCGenomics, 2010, 11:
262-273다.
[59] Luo HM, Sun C, Sun YZ 외. Panax 노토삼 뿌리의 전사체 분석을 통해 추정 triterpene saponin-생합성 유전자와 유전자 표지를 밝혀낸다 [J. BMC Genomics, 2011, 12:S5.
[60] 한 JY, 황 HS, Choi SW, et al. Cytochrome P450 CYP716A53v2 catalyzes the formation of protopanaxatriol에서 protopanaxadiol 동안 ginsenoside biosynthesis 파-낙스 인삼 [J]에서. 식물세포생리학, 2012, 53:1535-1545.
[61] Chen X, Xue Y, Liu JH 외. Panax 인삼털뿌리배양으로부터 glucosyltransferase의 정제 및 특성 (J. Pharm Biotechnol, 2009, 16:50-54.
[62] Yue CJ), 종제이 (Zhong JJ. UDPG:ginsenoside의 정제 및 특성 Rd glucosyltransferase from 정지 세포 of Panax noto삼 [J]. Proc Biochemistry, 2005, 40:3742-
3748.




