

 영어
영어 프랑스
프랑스 스페인
스페인 러시아
러시아 한국
한국 일본
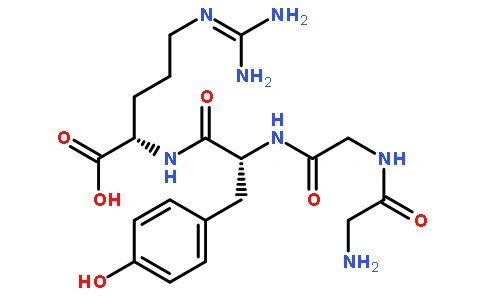
일본사료산업에서의 해피 (害皮)와 그 용도는?
Papain enzymes are primarily found in 이roots, stems, leaves, 그리고fruits 의papaya, with the highest concentrations in the 라 텍스의unripe fruits (Ye Qiteng etal., 1999; Zhao Yuanfan etal., 1999; Yi Yin et al., 2000). Due to their strong protein-degrading ability and the ability to hydrolyze amide bonds and ester bonds, papain enzymes are widely used in the pharmaceutical, food, textile, leather, feed, and dye industries (Wu Xianrong et al., 1988).
Papain 효소의 개요 1
파파 (papaya)에서 유래주스는 순수한 효소가 아니다.이러한 조효소에는 파파인뿐만 아니라 리소자임, 시스테인 프로테아제, 셀루라제, 글루카네이스, 글루타민, 저분자 티올 화합물 등이 함유되어 있다.파파야 라텍스에 함유된 시스테인계 효소는 등전점을 기준으로 크게 3가지로 분류할 수 있는데, 파파인 복합체 (papain complex, chymopapain), 리소자임 (lysozyme)이다.파파인은 9.55의 파이, 치모파파인은 10.10의 파이, 리소자임은 파이>11.0. 중성에 가까운 조건에서이 세 가지 효소는 케이션 교환 컬럼 크로마토그래피로 쉽게 분리할 수 있습니다.파파야 라텍스에서 파파인은 수용성 단백질의 약 10%, 키모파파인은 45%, 리소자임 (lysozyme)은 20%를 차지한다 (Ling Xinghan et al., 1998).
")
해독 (害毒)의 구조 2
2. 1 Papain
파파인 (Papain)은 최초로 발견되고 연구되었으며 널리 응용된 효소이다.1937년, Balls와 Lineweaver는 등급을 매긴 소금 강수법을 사용하여 신선한 파파야 주스에서 결정질 파파인을 추출했다.1970년에는 파파인의 염기서열을 규명하였고, 1971년에는 x 선 결정학을 이용하여 3차 구조를 규명하였다.
파파인은 상대분자량이 21,000 이며 212개의 아미노산 잔기로 구성된 하나의 폴리펩타이드 사슬로 이루어져 있다.1969년 Husain과 Towe는이 효소의 아미노산 서열을 보고하였다 (그림 1 참조).
파파인은 7개의 시스테인 잔기를 포함하고 있으며, 그 중 6개는 3 사슬 내에서 이황화결합을 형성하고, 나머지 자유시스테인 잔기는 그 중 하나이다필수 아미노산활성 중심의 잔류물입니다.Drenth 등은 파파인 분자의 3차 구조를 결정하였는데, 파파인 분자는 모양이 타원형이며 두 개의 도메인으로 구성되어 있고, 접합부에 좁은 홈이 있어 효소의 활성부위가 위치한다 (Chen Zizhen, 1978).
커드 파파인 2.2
파파인은 1941년 얀센과 볼스에 의해 처음 발견되고 추출되었으며 명명되었다 (얀센 외, 1941).papain의 구성은 비교적 복잡하다.1967년 쿠니미츠 등은 이온교환열을 이용하여 파페인을 분리하고 용출순서에 따라 두 종류로 분류하였다:파페인 A와 B;1982년 브로클허스트 등은 파파인이 4가지 성분으로 이루어져 있다고 보고하였다.1985년에는 극도로 느린 NaCl 구배 용출을 이용하여 5개의 활성 정점을 얻었다 (Kunimitsuet al., 1985;Brocklehurst et al., 1987)이다.
Silvia 등 (1989)은 papain의 4가지 하위유형에 대해 circular dichroism 분석을 실시하였다.현재 파파인은 서로 다른 아형으로 존재하며, 이들 사이에 아미노산 차이는 한두 가지에 불과하다는 것이 밝혀졌다 (버 틀et al., 1984).EMBL/GenBank/DDJB와 같은 데이터베이스의 자료에 따르면 적어도 9개의 아형이 있으며 각각 다른 유전자에 의해 부호화되어 있다.이 중 6개의 아형은 218개의 아미노산을 가진 폴리펩타이드 사슬로 구성되어 있고, 2개는 227개의 아미노산, 1개는 226개의 아미노산을 가지고 있다.9가지 아형의 아미노산 서열이 그림 2에 표시되어 있다.비교 결과는 이들 9개 아형들 중에서 제2 아형, 제3 아형 및 제 V 아형만이 특정 부위에서 개별 아미노산 잔기가 다른 아형들과 차이를 보이는 것으로 나타났다.
Maes et 알다.(1996)과 Azarkan et al. (1996)은 파파인의 3차원 구조를 규명하였다 (그림 3 참조).그림 3에서 보는 바와 같이 파파인의 폴리펩타이드 접힘은 크기는 같지만 모양이 다른 두 개의 도메인 (L 도메인, R 도메인)을 형성한다.L 도메인은 주로 α로 이루어 져-helices, R 도메인은 주로 anti-parallel로 이루어 져 있는 동안 β-sheets다.활성 사이트는이 두 도메인 간의 인터페이스에 있습니다.Papain에 속하는 α + β 클래스, 어디 Papain의 C-terminal 도메인은 완전 한 α-helical 구조, N-terminal 도메인을 완전히하는 동안 β-sheet 구조 (슐츠 et al., 1979).대조적으로, papain에 속하는 α/β 클래스, 보조 가들어 있는 구조로 더 α-helices와 β-sheets, 그리고 보조의 접 패턴 구조는 또한 다른 (Ssolis-Mendiola et al., 1992)이다.
파파인은 8개의 시스테인 잔기를 포함하고 있으며, 그 중 6개는 분자 내 이황화 결합을 형성하고, 나머지 2개의 활성 자유 설프히드릴기는 잔기 25와 117에 위치한다.그들의 상당한 거리와 다른 지역 때문에,이 두 잔기는 분자 내 이황화 결합을 형성할 수 없습니다.두 잔류물은 비슷한 활성을 가지고 있다.papain의 3차원 모델에서 Cys117은 분자 표면에 위치하여 유도체화 시약에 쉽게 접근할 수 있으며 완전하고 비가역적인 산화에 취약합니다.보존되는 Cys22 및 Cys56과 함께 촉매 Cys25는 S1 마이크로사이트를 형성한다.S1 마이크로 사이트의 비교적 큰"주머니"로 인해 효소&에는 거의 아무런 영향을 미치지 않는다#39, s 기질 특이성.그러나 67, 68, 69, 133, 157, 207위치에 잔류물로 구성된 S2 마이크로사이트는 효소에 영향을 미친다#39;s 기질 특이성 (Maes et al., 1996).
파파야 리소자임 2.3
1955년 스미스 등이 고립되었다순수한 결정 (結晶)파파야 라텍스.이후 Haward와 Glazer는이 효소의 구조와 특성을 규명하였다.파파인은 분자량이 2만 4000여개로 동물성 리소자임보다 높다.223개의 아미노산으로 구성되어 있으며, 아미노산 조성은 표 1에 나타내었다.파파인은 동물성 리소자임 (lysozyme)에 비해 프로일린 (proline), 티로신 (tyrosine), 페닐알라닌 (phenylalanine)이 현저히 높다.

n-말단의 아미노산 배열은 다음과 같다:glycine-isoleucine-serine-isoleucine.c-말단의 아미노산의 배열은 다음과 같다:세린-페닐알라닌-글리신.papain의 상위 구조는 원형 dichroism 분광법에 의해 결정되었으며, 단일 나선체가 전체의 약 30%를 차지하였다.분광측정과 화학시약반응을 보면 트립토판잔류물은 전적으로 효소분자내에 박혀있으며 n-아세틸글루코사민의 결합이나 활성반응에 참여하지 않는다.Papain lysozyme에는 8개의 시스테인 잔기가 포함되어 있으며, 그 중 4개는 이황화 결합을 형성하고, 4개는 자유 SH 그룹이다.하나의 SH 그룹은 효소&에 필수적이다#39;s 활성 사이트.papain lysozyme의 활성 부위 구조는 egg white 및 human lysozyme의 활성부위 구조와 상당히 다르다 (Funatsu et al., 1982).
해피 (害皮)의 3가지 속성
3.1 害와 害의 프로테아제 특성
파파인 프로테아제의 구조적 유사성은 성질의 중요한 유사성을 결정한다.papain과 papain protease는 모두 넓은 기질 특이성을 나타내는데, 대부분의 펩타이드 결합은 어느 정도 papain에 의해 가수분해될 수 있지만, 가수분해 속도는 여러 펩타이드 결합에 따라 크게 다르며, 일부는 3차까지 차이가 있다;많은 아미노산 유도체와 펩타이드가 기질의 역할을 할 수 있으며, 그 중에서도 아르기닌 유도체는 가수분해에 특히 민감하다.그러나 여러 가지 결속을 속박 (俗) 하는 것은 속박 (俗)보다 훨씬 빠르다.
Jansen et al. (1941)은 papain casein protease 가 papain의 절반 정도의 비율로 casein을 가수분해한다고 보고하였다.사용 되는 Ebara 및 Yasunobue papain 카 세인 프로 테 아제의 β 체인을 hydrolyze 산화 된 인슐린 (과 산화 칼륨 permanganate), enzyme&을 강조하#39;s는 산성 아미노산 잔기 및 방향족 아미노산 잔기를 포함하는 펩티드 결합을 가수 분해하는데 유리하다.라일은 인슐린의 A, B 사슬과 글루타민산을 함유한 다양한 길이의 다양한 펩타이드 분절들을 연구할 때, 글루타민산을 함유한 펩타이드 결합을 가수분해할 때 파파인의 우점에 주목하기도 했다.papain은 수용액 매체에 비해 유기용매 매체에서 더 넓은 기질 특이성을 나타낸다는 점에 주목할 필요가 있다 (진언소 등, 2000).
Papain and papain chymopapaincan also hydrolyze amide bonds and ester bonds (Ling Xinghan et al., 1998). They can catalyze the synthesis of oligopeptides but produce multiple byproducts, which may be due to the broad substrate specificity of the enzyme's S1 site (진언소 외, 2000).
papain과 papain protease의 최적 pH 값은 기질에 따라 다르다.casein을 기질로 사용한 Papain protease는 최적 반응온도가 80°C (pH 7.0) 이며 37°C에서 최적 pH 범위가 3-5이다.Michaelis 상수 (Km) 값은 1.25 g/l (pH 7.0, 반응온도 37°C에서)이다 (Zucker et al., 1985).파파인의 최적 pH 값은 7.0 이며 pH 안정도 4~9이다.효소 활성은 60°C 이하에서 안정적으로 유지되며 90°C까지 일부 활성을 유지합니다.높은 열 안정성과 좋은 안정성으로 인해 사료 가공에 적합합니다.
PCMD,chlorobenzyl mercuric 산성, 요오드 아세 트 산, 요오드 아세 트 산, 과산화수소, NEM, 그리고 Hg과 같은 중금속 ² ⁺, Ag ⁺, Cu ² ⁺, 그리고 Zn ² ⁺ 군사교류 효소 활동 억제 할 수 있다.두 개의 자유 설프히드릴기를 함유한 파파인은 산화제에 의해 산화되거나 중금속 이온과 결합함으로써 산화에 더욱 취약하여 불활성을 유발한다.그러나, 다양한 환원제 (시스테인, 티오글리콜산,글 루타 티 온, DTT 등)과 일부 금속 킬레이트제 (EDTA)는 활성 중심부의 시스테인 잔기를 감소시켜 효소 활성을 활성화시킬 수 있다 (등경 등, 2004).
papain lysozyme의 특성 3.2
소자는 고수 할 수 있는 효소 β-1, 4 N-acetylglucosamine과 N-acetylglucosamine 사이의 결합 입니다.파파인 리소자임은 등전점이 10.5인 알칼리성 단백질로 리스테리아 모노사이토제네스에 대한 계란 백자 리소자임의 활성은 3분의 1에 불과하며 최적 pH 4.5, 최적 이온강도는 0.04-0.07이다.세포벽을 분해할 때 동물 유래 lysozyme과 유사하게 행동할 때 환원 말단에서 N-acetylmuramic acid를 생성합니다.파파인은 키틴을 분해하는데 높은 활성을 나타낸다.예를들어, 부패하는 젤리 각 소에서 활동는 계란 흰자 소자의 10배, 그리고 활동에서 200번이 부패하는 (N-acetylglucosamine) ₄은 계란 흰자 소자이다.효소들의 산물 ' s의 저하 (N-acetylglucosamine) ₅은 (N-acetylglucosamine) ₃ 그리고 (N-acetylglucosamine) ₂.n-아세틸글루코사민은 자유 상태로 존재하지 않으며, 당전달 반응은 거의 관찰되지 않는다 (So et al., 2000).
효소활성 측정 4
Papain 활동 확인 4.1
파파인의 활동은 효소&를 측정하여 결정된다#39;s는 지정된 조건 (특정 온도 및 pH 값 등)에서 기질 단백질의 가수분해를 촉매하는 능력으로, 단백질의 가수분해로부터 생성되는 아미노산의 양이 효소 활성의 단위로 작용한다.같은 조건에서 산생되는 아미노산의 량이 많을수록 효소는 더 크다#39;s 촉매반응능력과 효소활성이 높을수록.
파파인 활성을 결정하는 방법으로는 casein을 기질로 사용하는 UV 분광광도법, 인도페놀 색도법, BAEE (benzoyl-L-arginine ethyl ester)의 세 가지가 있다.각 방법에는 고유한 특징이 있습니다.이UV 분광광도법casein을 기질로 이용하는 것은 조작이 간단하고, 비용이 적게 들고, 효소정제시 효소활성을 비교하는데 적합하며, papain 생성물 및 식품효소의 활성을 확인하는데도 적합하다.인도페놀 측색법은 수행하기가 번거롭지만 간단한 장비가 필요하여 자원이 한정된 지역에서 사용할 수 있다;BAEE 방법은 안정적이고 재현성이 있어 효소의 성질에 대한 이론적 연구와 시약효소의 효소활성 판정에 적합하다 (우상룡, 2005).파파인 활성을 결정하는 미국 약사 및 중국 의약품 기준에 명시된 방법은 casein을 기질로 사용하는 자외선 분광광도법 (Luo Yanshou, 2000;천더밍 외, 2004).
자외선을 사용 할 때 spectrophotometric 효소 활성을 탐지하는 방법, 활동의 단위로 정의 되는:테스트 조건에서 필요 한 효소의 양에서 트리 클 산성 수용성 물질의 양을을 해방 시키기 위해 카 세인 hydrolyzed 분당, 흡 광도 275 nm의 파장을에 있을 때 동일하 흡 광도의 1 μ g/ml-dopa,은 효소 활동 단위 중 하나이다.
이 방법을 사용할 때 몇 가지 문제가 주의를 요한다:이처럼 트리클로로아세트산 농도에 따라 다른 효소 활성 결과가 나온다.② 다른 기원의 Casein은 papain 활동에 현저한 차이를 나타냄;수입 카세인은 용해도가 더 좋고 효소 활성이 더 높습니다.다시 재결정은 최적의 pH 조건에서 실시되어야 한다 (Zhao Yuanfan et al., 1999).
papain 응고 단백질 분해효소 활성 측정 4.2
파파인 응고 단백질 분해효소의 가수분해 활성을 확인하는 방법은 casein을 기질로 하는 UV 분광광도법을 사용한다.활동이 효소 또한 갖고 있 응고 되게 papain과 비교 했을 때, 응고 활동의 단위로 정의 되는 응고하는 데 필요 한 효소의 양을 10% 1 ml의 우유를 포함하는 (0. 01 mol/l CaCl ₂) 40분 이내에, 어느로 정의 되는 효소 활동 단위, 즉 1 Soxhlet 단위 (수애)이다.상대활성도 (relative activity, RU)는 다양한 요인의 영향을 나타내는 데 사용된다 (Arima et al., 1967).
4. 3 Papain lysozyme 활성 판정
리소자임의 활성을 결정하는 방법에는 일반적으로 3가지가 있다.
이처럼 세포벽을 기질로 사용하는 효소활성은 작용 전후의 탁도의 변화로 나타난다.
② Listeria monocytogenes의 배양액을 기질로 하여 작용 전후의 탁도의 변화로 효소활성을 나타낸다.
상기 두 가지 방법은 고체-액체상 반응을 포함하기 때문에 반응속도를 정확하게 측정하기 어렵다.따라서, 일부 연구자들은 사용하여 균질한 기질을 준비했습니다수용성 다당류효소 활성을 확인하기 위해 hexane diol chitin.
다시 한 번 분광광도법:450 nm에서 일정량의 동결건조된 박테리아 분말 또는 해동된 용해불산 박테리아 유지를 일정량의 인산완충된 용액에 녹여 약 1.3의 흡광도 값을 얻는다.이 기질 용액과 표준 효소 용액은 수조에서 25°C에서 배양된다.수조에 있는 1 cm 큐벳에 기질 2.5 ml를 넣고 효소용액 0.5 ml를 넣고 시작 타이밍, 1분에 판독 E1, 2분에 판독 E2를 기록하고 수식을 이용하여 효소활성을 계산한다.다른 표준 효소 솔루션에 대응하는 다양 한 효소 가 활동 있는 다른 Δ E 값을, 그리고 효소는 데 사용 되는 표준 효소의 활동을 계산 할 수이 러한 Δ E 값에 기초 하여, 그렇게 함 으로써 확인의 정확도를 탐지 한다. (2004년 장 [용],)
害의 고정화 5
고정화 효소는 1960년대에 개발된 신기술이다.고정화란 자유세포 또는 효소를 물리적 또는 화학적 방법을 사용하여 고체 불용성 운반체와 결합하여 활성을 유지하고 반복적 사용이 가능하도록 하는 과정을 말한다.papain의 활용 효율성을 향상시키고 생산 비용을 절감하기 위해 전세계 연구자들은 papain의 고정화 및 적용에 대한 광범위한 연구를 수행했습니다.
1961년, j. Cebray 성공적으로 움직이 papain polyamino 한 산성에 캐리어와 그것을 이용 해 hydrolyze γ의 파편-globulin다.1977년, 미국의 J.W. 핀리는 키틴에 파파인을 고정시키기 위하여 글루타알데히드교반법을 채용하였고 맥주생산과정에 사용하여 좋은 해명효과를 거두었다.1978년 프랑스의 P. Monsan 등이 aminoalkylated microporous glass를 운반체로 사용하여 표면에 papain을 교차 연결하였으며, 맥주해명에 적용하기도 하였다 (Ling Xinghan et al., 1998).
Xu Fengcai et al. (1992)은 papain을 고정화하기 위한 운반체로 사탕수수 배지 셀룰로오스와 나일론을 사용하였으며, 고정화 효소의 효소적 특성을 규명하고, 이를 맥주 해명에 적용하였다.이홍 등 (2001)은 키토산 미립구를 이용하여 파파인을 고정화하고, 고정화 효소의 효소적 특성을 연구하였으며, 이를 티로신 생성을 위한 카제인 가수분해에 적용하였다.또한 저자들은 누에로부터 실크섬유를 이용하여 파파인 고정화, 고정화 효소의 효소적 특성을 조사하여 tyrosine 생성을 위한 casein 가수분해에 적용하였으며, packed-bed 반응기에서 실크-고정화 파파인과 그 적용방법, 모두 만족할 만한 결과를 얻었으며 (Chen Fangyan et al., 2004, 2005a, 2005b), 해당 고정화 파파인 생성물을 얻었다.

6 사료산업에서의 papain 적용
식품 가공 중,의 많은 양단백질 부산물동물의 깃털, 도살 후 동물의 피, 생선 손질, 생선 가공에서 나온 생선 머리 등 생성됩니다.이런 단백질은 직접 사료로 갈면 동물이 소화, 흡수하기 어렵다.그것들을 버리는 것은 자원을 낭비할 뿐만 아니라 환경을 오염시킵니다.단백질분해효소를 리용하여 이런 단백질을 가용성 소분자단백질과 아미노산으로 가수분해하면 동물이 쉽게 소화, 흡수할수 있게 된다.이를 통해 저가의 고품질 단백질 사료 개발이 가능할 뿐만 아니라 사료이용효율을 향상시키고 사료비를 절감시킬 수 있다 (노성민 등, 2001;탄아이주안 외, 1998;양평 외, 2008).현재 동물성 단백질 효소는 가수분해하기에는 너무 비싸다;미생물 발효 방법은에 의해 오염되기 쉽습니다 다른 박테리아 및 독소를 포함 할 수 있습니다;그러나 파파인은 활성이 강하고 열성이 높으며 안정성도 좋아 사료가공에 적합하다.
害身 [명사] 【 문어 】 害身 paain가축 식이에 첨가하는 사료 첨가제, 사료 소화에 도움을 주고, 사료 효율을 향상시키며, 사료 섭취량을 줄이고, 가축의 성장 속도를 향상시키며, 우유 생산, 우유 품질, 젖소의 유방염 예방에 상당한 영향을 미친다.
Zhang Qing 등 (1996)은 새우 사료에 0.3% papain을 첨가하여 생산 중 papain 활성의 변화와 양식장에서의 효과를 연구하였다.그들은 새우 사료 생산 조건 (85°C, 45분)에서 papain 활성 손실이 심각하다는 것을 발견했다.그러나 가금류 사료 생산 조건 (온도 75°C) 에서는 protease 활성이 상당히 잘 보존되었다.그러므로 골반염조건에서 대부분의 효소활성이 보존될수 있는 온도는 75°C 정도이다.새우양식에서는 일정한 생장촉진효과를 증명하여 생산량을 5% 증가시켰다.
Binshiyu 등 (1996)은 돼지를 재배하는 식이에 0.1% papain을 첨가하여 일일 체중증가와 사료전환율을 향상시켰으며, 10~20 kg의 돼지에서 가장 큰 효과가 관찰되었다 (p<0. 01)이다.이 결과는 돼지를 기르는 초기 성장기에 파파인을 첨가하는 것이 가장 적절하며, 중기 성장기에도 적당량을 사용할 수 있음을 보여준다.

허팅 등 (1992)은 육류형 병아리들에게 파파인 (사료 40,000단위/kg)으로 보충한 식단을 먹였다.그 결과 그룹 간 평균 체중 증가량에는 유의한 차이가 없었지만supplemented with papain (P > 0.05), feed consumption was slightly lower in all experimental groups compared to the control group. Additionally, the use of FS 단백질feed (fermented blood meal) in the diet was more effective than fish meal. 이reason may be that FS protein feed contains not only animal blood but also various plant fiber-rich cake and meal carriers, which may allow other enzymes in papain to exert their effects.
7 결론
파파인은 순수 천연물로 강력한 단백질 분해 가수분해 및 합성 능력과 응고, 립분해, 박테리아 분해 활성을 가지고 있어 다양한 효능이 있다.현재 파파인의 세계 적용 비율은 다음과 같습니다:양조 및 음료 산업에서 75%, 육류 가공 산업에서 10%, 생선 가공 산업에서 5%, 제약 산업에서 3%, 사료 가공 산업에서 5%, 기타 분야에서 2%입니다.사료산업에서 papain의 적용 비율이 상대적으로 낮은 것으로 나타나 향후 상당한 발전 가능성이 있음을 알 수 있다.그 이유는 다음을 포함할 수 있습니다:사료 처리 중 효소의 상대적으로 높은 비용, 응용 프로그램을 제한하는 효소 활성의 상당한 손실;또는 사료첨가제로서 효소는 사료공급과정에서 과학적으로 사용되지 않는다.따라서 papain의 활동을보다 효과적이고 지속가능하게 향상시키고 유지하는 방법;그리고이 효소를 다양한 동물 종과 다양한 먹이 공급 단계에서 정확하고 합리적으로 사용하여 효능을 극대화하는 방법은 향후 연구 및 개발 노력이 필요한 가치가 있는 핵심 분야입니다.
참조
[1]Ye Qiteng, Chen Qiang.害用 [명사] 해용 (害用).광시 열대작물 과학기술, 1999(4):34-35.
[2] 자오위안판 (Zhao Yuanfan), 딩렌캉 (Ding Renquan).Papain [명] 가공기술과 응용 (製造).연난사범대학 논문집 1999, 19(5):46-48.
[3] 이인, 탄아이후안, 류닝.해파 (害派)의 제작과정에 관한 연구 (硏究)귀농과학, 2000, 28(5):24-25.
[4] 우샹룽, 주리칸.Papain다 [J다]북경농업대학 논문집, 1988, 14(1):13-17.
[5] 링싱건, 우샹룽.Papain과 Papaya 재배 [M.북경:중국농업신문 1998:104.
[6] 천자이젠.식품효소학 [M.대만:Fuwen 출판사, 1978:142.
[7] Kunimitsu K, Yasunoba K T, Chymopapain 나는 V. 이 chromato-그래픽 fractionation of 부분적으로 정화 chymopapain and the 결정질 chymopapain [J]의 특성.바이오케미카 et Bio- physica Acta, 1985, 828(2):413-417.
브로 클 허스트 [8] K, Willenbrock F, 살리 E. 시스테인 단백효소 [J.신종합생화학, 1987,16:139-158.
[9] 실비 S M, Rafael Z L, A.Rojo-Dominguez 외.원형 dichroism [J]에 의해 나타난 chymopapain 형태의 구조적 시미-larity. 생화학 학회지, 1989, 257:183-186.
버 틀 [10] D J, 바렛 한 J. Chymopapain 크로마토그래피 퓨리피카-션 및 면역학적 특성 [J]다. 생화 학적 저널, 1984년, 223 (1):81-88.
[11] Maes D, Bouckaert J, Poortmans F, et al. 구조 chymopa의-1.7A 해상도의 고통 [J].생화학, 1996년, 35:16292-16298.
[12] Azarkan M, Dominique M, Julie B 외.Thiol pegylation facili-chymopapain의 tates 정제로 1.4A 해상도의 회절 연구를 유도합니다 [J]다.한국크로마토그래피학회지 A, 1996, 749:69-72.
[13] 슐츠 G E, 측정 할. 의 원칙 protein 구조 [M]다. 뉴욕:SpringerVerlag, 1979:44-107.
[14]Ssolis-Mendiola S,Arroyo-Reyan A, Hernandez-Arana A. circu-lar dichroism of cysteine proteinases from 파파야latex-their polypeptide chain의 접힘의 차이를 보여주는 증거.biochem [J].Biochemicaet BiophysicaActa, 1992, 1118:288-292 쪽.
[15] 후나츠, 카츠토시, 츠루, 다이스케.번역 Li Xingfu.소자 [M]이다.제남:산동과학기술출판사, 1982:74-76.
[16]Jansen E F, Balls A K. Chymopapain:에서 나온 새로운 결정질 단백질 분해효소 papaya latex [J]다. 이 저널 of 생물학적 화학, 1941년 137:459-460다.
소진언 (Jin-Eon So), 신종식 (Jong-Shik Shin), 김병기 (Byung-Gee Kim)프로테아제 촉매 트리펩타이드 (RGD) 합성 [J.효소와 미생물기술, 2000,26:108.
주 커 [18]. S, Buttle D J. The proteolytic 활동 chymopapain, papain, 파파야 단백분해효소 Ⅲ [J]다. Biochemica et Biophysica Acta, 1985, 828(2):196-204.
[19] 등징, 자오수진.부제:Enzymatic properties of papaya coagulating proteinase [J].아미노산과 생물자원, 2004, 26(2):18-20.
[20] 그래서 신제이 (J E, Shin) J S, Kim B G. Protease 가 촉매되었다 리펩타이드 (RGD) 합성 [J].효소와 미생물기술, 2000, 26(2):108-114.
[21] Wu, X. R. papain [J] 개발 및 응용.「 중국농업대학 논문집 」, 2005, 10(6):11-15.
[22] Luo, Y. X. papain activity 결정방법에 관한 연구.중국약학학회지 2000, 35(8):556-558.
[23] 천더메이, 푸슈후안.Papain Enzyme Activity Assay 방법의 실제 생산에서의 적용 가능성 [J].광동제약대학 논문집 2004, 20(3):244-245.
[24] Arima K, Shinjiro I, Gakuzo T. Milk clotting enzyme from microorganisms, 제1 부, Screening test and identification of potent fungus [J.농업생물학 및 화학, 1967, 31(5):540-545.
[25] 장용.라이소자임과 식품산업에서의 응용 [J.곡물 가공 및 식품 기계, 2004(3):64-65.
[26] 쑤펑카이, 장웨이, 뤄강위 등.사탕수수 배지 셀룰로오스에 대한 Papain의 고정화 및 그 적용에 관한 연구 (J.남중농업대학 논문집, 1992, 13(1):53-59.
[27] 쑤펑카이, 리밍키.Nylon에 Papain의 고정화와 그 응용에 관한 연구 (J.생화학, 1992, 8(3):302-306.
[28] 이홍, 왕웨이준, 서풍케이.Chitosan microsphere-immobilization Papain의 특성에 관한 연구 (J.남중농업대학 논문집 2001, 22(2):56-58.
[29] 천팡옌, 지핑슝.누에실크 피브로인에 대한 Papain의 고정화에 관한 연구 (J.남중농업대학 논문집 2004, 25(3):83-86.
[30] 천팡옌, 지핑슝.실크 피브로인에 고정화된 Papain의 특성에 관한 연구 (Study on the Characteristics of Papain immobilization on Silk Fibroin [J])전남농업대학 논문집, 2005a, 26(4):81-83.
[31] 천팡옌, 지핑슝.실크 피브린 고정화 파파인의 충전층 반응기 및 그 응용에 관한 연구.설화과학, 2005b, 31(3):286-289.
[32] 류시민, 탄아이 후안.돼지 혈식단백질에 대한 papain의 가수분해 작용 [J.귀농과학, 2001, 29(4):6-7.
[33] 탄아이 후안, 양송, 시앙 랑타오.사료단백질의 파파인 가수분해를 위한 최적 조건 (J.한국산농생물학회지, 1998, 17(2):106-109.
[34] 양평, 하영준, 판웨이캉.tilapia 부산물에 대한 papain의 가수분해 효과 (J.수산과학, 2008, 27(6):290-292.
[35] 장칭, 리주쟈오, 천캉글 등.새우사료에 papain의 첨가에 관한 연구 (J.사료산업, 1996, 17(5):8-10.
[36] 빈시유, 판시종.돼지를 기르는 식단에서의 papain 적용 [J.곡물 및 사료산업, 1996, 17(7):24-25.
[37] 셰팅, 량린, 판쑤이화 등이 있다.육계 병아리의 식이에 해파인을 첨가하는 효과 (effect of adding papain to the diet of broiler chicks)광동성 축산수의학회지 1992(2):9-11.




