

 영어
영어 프랑스
프랑스 스페인
스페인 러시아
러시아 한국
한국 일본
일본은행추출물 은행골산을 어떻게 합성하는가?
Ginkgo bilob한L. is 이oldest living tree species on 이planet그리고is known as 이“golden living fossil”. As a 독특 한economic gymnosperm 에서China, it has multiple values as food (white fruit), medicine (leaves 그리고white fruit) 그리고ornamental plants. Ginkgo nuts (seeds) are a famous health-promoting dried fruit 에서China. As a food 와medicinal properties with high nutritional value, the ginkgolides contained 에서it are potential 활성ingredients against the 2019-nCoV virus[1]. Ginkgo biloba leaves are rich 에서flavonoids 그리고lactones. Extraction 의Ginkgo biloba (EGB) with acetone-water as the extracting agent can further enrich the two active substances 을effectively treat cardiovascular 그리고cerebrovascular diseases [2]. Ginkgo biloba is a world-famous ornamental tree for landscaping with its unique leaf shape, golden leaf color 그리고upright tree shape. At present, ginkgo has been extensively developed 에서the pharmaceutical, food, cosmetics, bonsai and timber industries, and is one 의the key economic tree species for the current and future revitalization 의the rural economy and the construction 의a beautiful China.
1970년대부터 차이나'의 은행나무 자원은 급속히 증가하였고, 부존자원은 세계&의 80% 이상을 차지하고 있다#총 39; s다.은행나무는 하이난, 헤이룽장, 내몽고를 제외한 모든 성 (다른 곳에서 도입한 성 포함)에 분포한다.현재 중국의 은행나무 재배 면적은 33만 3300 hm2 이상이며, 연간 은행나무 견과류 1만 1000여 톤, 말린 녹색 잎 1만 톤을 생산하고 있다.은행재배 1차 산업의 연간 생산액은 약 20억 위안, 은행가공 및 의약품 생산 (은행잎약)의 2차 산업은 100억 위안 이상의 가치가 있다.

하지만, 매년 약 3만 톤의 외부 씨앗 코트가 생산으로 인해 버려집니다.장신희 등 3명은 겉씨저고리에는 다음과 같은 활성물질이 함유되여있음을 보여주었다ginkgolic acid, flavonoids, terpene lactones and polysaccharides. Itokawa et al. [4] showed that ginkgolic acid is another important active substance 에서ginkgo biloba, 에서addition 을ginkgolide and ginkgoflavone. It has bactericidal, bacteriostatic, anti-inflammatory, anti-viral, insect repellent and insecticidal effects, and is 의great value for development. Therefore, the large amount 의discarded outer seed coats not only pollutes the environment but also causes a waste of resources.
1970년대에 Gellerman et al. (5-7)은 은행잎, 미성숙한 종자 및 성숙한 겉씨코트 (outer seed coats)에서 phenolic acid ginkgolide를 검출하였다.이후 정량측정을 통해 ginkgolide는 성숙한 겉씨 외투에 농축되어 있음을 알 수 있었다.왕지 등 [8]과 이홍청 등 [9]은 질량분석법을 이용하여 은행산성 추출물에서 4개의 은행산과 2개의 은행산을 확인하였다.현대의학연구에 의하면 은행골질은 일정한 알레르기성 독성과 세포독성을 갖고있으며 세균과 진균의 생장에 저항할수 있고 해충을 죽이는 효과가 있다는것이 증명되였다.
밭재배 실험을 통해 은행골류가 생농약으로 개발될 수 있음이 입증되었으며, 향후 의약과 화장품 등 다양한 분야에 활용될 수 있을 것으로 보인다.그러나 은행플라보노이드, 테르펜락톤과 같은 명확한 약리학적 활성을 가진 물질에 비해 은행산의 생합성 경로, 조절 기전 및 유도체 효능 등에 대한 연구가 깊이 있게 이루어지지 않아 은행산의 합성을 억제하기 위한 분자생물학 기술과 은행산의 높은 산출량을 생산하기 위해 발효시키는 공학적 응용에 대한 연구 및 개발이 늦어지고 있으며,그리하여 은행업의 더 많은 발전을 저애하고있습니다.

이러한 점을 감안하여 필자는 최근 은행골라이드의 생물학적 활성에 관한 기초 연구를 요약하고, 지방산 합성과 폴리케타이드 합성으로 구성된 은행골라이드의 합성 경로를 상세히 설명하며, 합성에 관여하는 주요 효소들의 촉매 메커니즘을 요약하고, 앞으로의 연구 방향을 전망하여 차후 은행골라이드에 관한 연구에 참고가 되고 이론적 뒷받침이 되는 것을 목적으로 한다.
은행나무의 생물학적 활성 1
은행골산은 페놀성 지질군에 속하며, 알킬페놀산, 알킬레조시놀산, 아나카르다산, 알킬카테콜스 [10] 등 크게 네 가지로 나뉜다.그들은 식물의 2차 대사산물의 한 종류이다.부동한 종류의 식물은 독특한 페놀성물질을 함유할수 있다.이들의 주요 생리기능은 생식 및 무생물 스트레스에 저항하는 것이다.
1.1 Ginkgolide 조성과 물리적, 화학적 성질
은행나무 (Ginkgolides)는 은행나무 (ginkgo biloba)의 중요한 2차 대사산물로 옻나무과에 속한다.은행산 (ginkgolic acid), 은행골 (ginkgol), 빌로볼 (bilobol) [11]의 세 가지 성분을 포함하고 있다.은행산은 측쇄 길이가 13, 15, 또는 17 이고 측쇄 이중결합수가 0 내지 2인 2-히드록시-6-알케닐 (알킬닐) 벤조산이다 (도 1a).ginkgolic acid B, ginkgolic acid A, ginkgolic acid, heptadec-1-enylginkgolic acid, heptadec-1-dienylginkgolic acid 등 총 5 종류가 분리 및 확인되었다.은행골라이드는 측쇄의 길이가 15 또는 17 이고 측쇄의 이중결합 개수가 1인 3 알케닐페놀이며, 총 2가지 종류가 있다;은행산은 측쇄의 길이가 15 또는 17 이고 측쇄의 이중결합수가 2인 5-alkenylresorcinols 이며 (Fig. 1b), 총 두 종류가 있다.
Plants such as 제라늄(Pelargonium hortorum) in the Geraniaceae family [12] and sumac (Anacardium occidentale) in the Anacardiaceae family [13] also contain urushiolic acids. The alkyl/alkenyl side chain lengths and the number of unsaturated bonds, and so on, but the chemical structure is similar 을that of ginkgolides. Ginkgolic 산are the main substances in ginkgolides extract, accounting for 90% of the entire acidic extract. The proportions of the five ginkgolic acids are different, mainly including ginkgolic acid (50% content), followed 에 의해heptadec-1-enyl ginkgolic acid (22%) and ginkgolic acid (20%) [14] (Figure 2). With the development of science and technology, ginkgo acid has been detected in ginkgo leaves, outer seed coats and kernels. The total ginkgo acid content of ginkgo leaves from different varieties (strains) is about 14.5765-23.6813 mg/g [15], and the total ginkgo acid content of mature kernels is about 0.11 mg/g [14]. and the total ginkgolic acid content in the outer seed coat can reach 28.78 mg/g [16].

The melting point of ginkgolic acid is 41-42°C. At room temperature, pure ginkgolic acid is oily or powdery in appearance, and is poorly soluble in polar solvents such as water or ethanol, but easily soluble in non-polar solvents such as light petroleum ether. It precipitates as crystals in saturated petroleum ether solvent [17]. In solution, the hydroxyl and carboxyl groups on the phenolic acid benzene ring ionize to produce a weak acid, which can undergo esterification and saponification reactions. Ginkgolic acid that has undergone esterification or saponification is easier to extract and separate, achieving the effect of purification. The melting point of ginkgolic acid is about 136 °C and the boiling point is about 500 °C. At 200 °C, the carboxyl group on the phenolic acid benzene ring will undergo a decarboxylation reaction to release CO2, which can be separated using the temperature gradient method. Therefore, according to physical and chemical property research, in the processing of ginkgo products, methods such as “hot air cooking”, “ultrasonic-assisted extraction of ginkgo phenolic acid with resin adsorption” and “compatibility of Chinese medicinal herbs” are often used to remove phenolic acids to achieve the purpose of detoxification. Chen et al. [18] latest 연구results show that the laccase immobilized on a new 유형of electrospun nanofiber felt can catalyze the degradation of ginkgo phenolic acid. However, the above-mentioned methods for dephenolic acid treatment have disadvantages such as high cost and difficulty of operation, and are not suitable for large-scale detoxification.
은행나무의 생물학적 활성 1.2
은행골라이드는 항균, 살충제, 알레르기성, 세포독성 및 항암효과가 있다.그들의 다양한 생물학적 활동은 농업 생산, 의료 및 다른 분야에서 이용될 수 있습니다.
항균 효과 1.2.1
은행산은 6-alkylsalicylic acid로 다양한 항균성을 가지고 있습니다.Bacillus subtilis, Escherichia coli, Staphylococcus aureus, Shigella dysenteriae, Pseudomonas aeruginosa, 그리고 다양한 그람 음성균과 그람 양성균에 대한 억제 효과가 있다 [19-20].이 산성 추출제는 벼잎마름곰팡이, 토마토시름곰팡이, 사과무트라코곰팡이, 옥수수잎점곰팡이, 붉은곰팡이 등에 억제 효과가 있으며, 항균 효과는 일부 항진균제에 필적한다 [21].서이천 등 [22]은 0.1%의 은행산 (15:1)은 균류를 억제하는데 92%의 유효율을 보였지만 0.5%의 클로트리마졸은 68%의 효과만을 보였다.Muroi 등 [23]은 은행산이 메티실린 내성 황색포도상구균을 사멸하는 데 표준 항균제와 시너지 효과를 보였으며, 48시간 내에 복합제의 살균 활성이 단일 제보다 최소 100배 이상 높은 것으로 나타났다.
살충효과 1.2.2
Ginkgolide acid has been shown to have a significant killing effect on aphids, grubs, cabbage moths, spider mites, mulberry ticks, rice borer and other chewing mouthparts insects [20]. In a test to 제어aphids, Shi Qitian [24] found that the killing effect of ginkgolide acid extract was comparable to that of the pesticide imidacloprid. Deng Yecheng et al. [25] used different polarities of exocarp extracts in contact toxicity tests and found that they had a strong killing effect on the brown planthopper, peach aphid, red spider and cabbage butterfly larvae.

1.2.3 알레르기 효과
In 1934, Hill et al. [26] reported that an active substance in ginkgo has a strong erosive effect on the skin. In 1969, Gellermen isolated lacquer phenolic acids from cashew nuts and ginkgo seeds, respectively, and pointed out that they have a strong sensitizing effect on the skin [5]. Cheng Liang et al. [27] reported that ginkgolic acid (C15:1) is metabolized to ginkgolide, which is further oxidized to form catechol, causing an allergic reaction. Vincieri et al. [28] showed that ginkgolic acid can inhibit the activity of various dehydrogenases in glucose metabolism. Ahlemeyer et al. [29] found that ginkgo acid has a competitive inhibitory effect on glycerol-3-phosphate dehydrogenase. Some scholars speculate that ginkgolic acid has a bipolar nature (hydrophilic and lipophilic) and can inhibit the activity of enzymes related to the body or organs, there에 의해affecting metabolism and causing allergies [30].
1.2.4 Cytotoxicity
Ginkgolide는 친수성군과 지방성군을 모두 가지고 있어 세포막을 가로질러 세포막에 결합하여 세포사를 유발할 수 있다.Al-Yahya 등 31)은 은행산이 함유된 은행추출물을 이용하여 수쥐 (male rats)를 대상으로 독성실험을 진행하였고, 그 결과이 추출물이 생식 세포와 같은 염색체 손실을 유발하여 쥐의 정상적인 생식 기능에 영향을 미칠 수 있음을 보여주었다.Ahlemeyer et al. [29]은 은행골산이 신경독성을 가지고 있으며 병아리 신경모세포종 세포를 죽게 할 수 있다는 것을 발견했다.최근 연구자들은 다양한 은행산의 독성에 대해 별도의 연구를 진행했다.Jiang 등 [32]은 은행산 (C15:1)이 퓨린 대사 장애를 유도함으로써 산화 스트레스를 유발하고 쥐 간세포에 손상을 줄 수 있다는 것을 발견했다.Yao 등 33)은 헵타데실리덴 은행산 (heptadecylidene ginkgo acid, C17:1)이 HepG2세포에 미치는 독성은 시간 및 용량에 정례 하며, CYP1A 및 CYP3A 대사를 매개로 하여 세포독성을 강화시킨다는 것을 밝혔다.연구 결과, ginkgolides 중에서 ginkgolic acid (C13:0), ginkgolic acid (C15:1), heptadecylidene ginkgo acid (C17:1) 가 독성이 더 강하고, 그 외 phenolic acid 가 세 가지 은행산과 결합하여 작용하여 세포독성을 증가시키는 것으로 나타났다.
항암효과 1.2.5
은행골라이드는 세포독성을 가지고 있고 항암 역할을 할 수 있다.Qiao 등 [34]은 은행산이 adenosine monophosphate-activated 단백질kinase (AMPK)의 활성화를 유도하여 증식과 이동을 억제할 수 있음을 발견하였다.Liang 등 [35]은 은행산이 ros를 조절하는 STAT3/JAK2 신호전달 경로를 억제함으로써 위암세포의 성장을 억제할 수 있음을 발견하였다.류 등 [36]은 은행산이 세포분열 G0/G1단계에서 대장암세포의 전이를 차단하여 암세포를 사망에 이르게 할 수 있음을 보여주었다.체외세포 실험 결과 은행산은 암세포의 증식과 이동을 억제하고 관련 효소를 활성화해 세포사멸을 촉진하는 것으로 나타났다.그리고 향후 종양 치료를 위한 보조 항암제가 될 수도 있다.
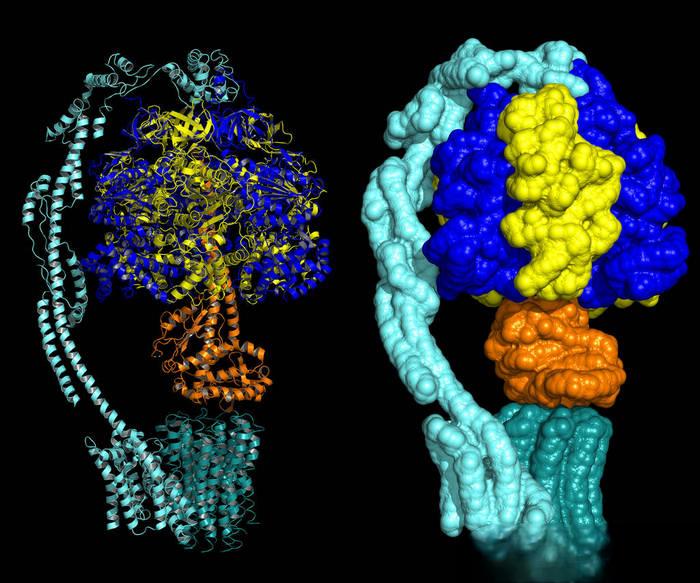
2 Ginkgolide 생합성
In the 1970s, Gellerman from the University of Minnesota in the United States used the 14C labeling method to conduct a preliminary exploration of the ginkgolide acid synthesis pathway. In the 1990s, Walters et al. [37] used gas-liquid chromatography (GLC trapping) to analyze labeled monomethyl esters extracted from liquid chromatography (HPLC), and determined the specific synthetic steps of 우라시올 (2-히드록시-6-alk (en)yl-benzoic acid)을 제라늄에 함유.Singhal [38]과 Narnoliya 등 [39]의 합성에 있어 주요 효소의 종류와 기능을 추가로 조사하였다 우 루시 올다.
2.1 Ginkgolide biosynthetic pathway를 사용한다
겔러만 (Gellerman) 등 (6-7)은 은행골라이드의 방향족 고리와 긴 사슬 알킬/알케닐 그룹 (alkyl/alkenyl group)이 단계적으로 합성된다고 믿고 있으며, 실험 추론 결과, 세 부분으로 나누어진다:이처럼 말로닐-coa와 아세틸-coa 형태가 없는 팔미토일-coa와 지방산 합성을 통한 올레오일-coa;② 긴 사슬의 acyl-CoA를 합성하여 polyketide 합성을 통해 ginkgolic acid를 만든다;다시 은행골산은 벤젠고리의 카르복실기를 잃고 산화되여 은행골과 같은 물질로 환원된다.
제라늄 (Pelargonium hortorum)은 제라니아과에 속하는 식물이다.이것의 삼색기는 시키믹산 (은행산의 동형체)만 분비하는데, 시키믹산 합성 경로를 연구하는 데 가장 좋은 종이다.야생형 제라늄은 포화 부사슬을 가진 시키믹산을 분비하고, 살충형 제라늄은 단일 불포화 부사슬을 가진 시키믹산을 분비한다 (C15:1, C17:1).연구 결과, 제라늄의 불포화 우루시올산은 이중결합의 위치 차이를 제외하고는 두 가지 우루시올산 (ginkgolic acid, C15:1)과 분자구조가 동일한 것으로 밝혀졌다.Hesk 등 12)은 야생형과 살충성 제라늄의 차별적 유전자와 대사산물을 비교하고, 우루시올 합성의 분자적 기전을 밝혔다.연구자들은 RNA-seq를 사용하는 제라늄을 구성하는 기술 cDNA transcriptome 데이터베이스, 될 수 있는하고의 핵심 유전자 합성을 식별를 Polyketide Synthases Ⅲ (PKS Ⅲ) 유전자와 Stearyl-ACP Desaturase (슬 픈)을 통해 유전 자가 유전자 주석 비교 및 차등 분석 표현이다.ginkgolide 합성의 조절 경로를 ginkgolic acid (15:1)와 heptadecylideneginkgolic acid (17:1)를 예로 사용하여 설명한다.

2.1.1 알킬 사이드 체인의 합성
ginkgolides의 합성은 먼저 palmitoleic acid와 oleic acid의 합성, 즉 alkyl side chain의 합성에 기초한다 (그림 3). 저장된 sucrose를 phosphoenolpyruvate로 발효, pyruvate kinase (피루브산 키나제)를 세포질에서 가소물로 형성시키기 위한 알로스테릭 변화, acetyl-CoA를 형성하기 위한 pyruvate dehydrogenase (피루브산 탈수소효소, PDH)의 촉매 작용은 지방산에 대한 시작 2C 분자를 제공한다.아세틸-coa는 아세틸-coa carboxylase (Ace-tyl-CoA carboxylase, ACC)에 의해 촉매되어 말로닐-coa를 형성한다.Transacylase (TA)는 CoA 분자를 acyl carrier protein (ACP)으로 대체하여 malonyl-ACP를 형성한다.Malonyl-ACP acetyl-ACP 되기 카르 그룹,을 잃고 하여 주기에서은 축약 된 β-Ketoacyl-ACP synthase 3세 (KAS III)의 시작을 형성 하기 위해 물질 4C 번 acyl-ACP다.
그럼 4C acyl-ACP은 반복적으로 축약 된 β에 의해-Ketoacyl-ACP synthase palmitoyl-ACP을 형성 하기 위해나는 (KAS I) (C16:0 ACP), 그리고 나서 dehydrogenated 되는 Δ 9-stearoyl-ACP desaturase (슬 픈) palmitoleic-ACP를 형성 하기 위해 (9 Δ C16:1 ACP)이다.슬 픈)은 dehydrogenated을 palmitoleic acid-ACP (9 Δ C16:1 ACP), 그리고에 의해 촉발시 ketoacyl synthase II (β-Ketoacyl-ACP synthase Ⅱ, KAS Ⅱ)를 형성 하기 위해 oleic acid-ACP (Δ 11 C18:1 ACP)다.가소물 중의 Palmitoyl-ACP와 oleoyl-ACP는 각각 티오에스터제 A (TE/FatA)와 티오에스터제 B (TE/FatB)에 의해 탈실화되어 자유 불포화 지방산 [40]을 형성한다.마침내, 자유 지방산으로 전환은 palmitoleyl-CoA (9 Δ C16:1CoA)와 oleoyl-CoA (Δ 11 C18:1CoA)에 의해 acyl-CoA synthase (ACS)의 외부 막에 plastid, 그리고 세포 질로 이송을 두 가지 중요 한 ginkgolic 산은의 전신이 되었다.
그러나 은행골산과 제라닐제라닐의 지방산 합성 경로는 다르다.ginkgolic 산의 합성에 대한 선임자들 (C15:1)와 heptadecylideneginkolic 산성 (C17:1)은 palmitoleic 산성 (9 Δ C16:1)와 oleic 산성 (Δ 11 C18:1), 2의 선임자들을 합성하는 동안 monoenyl 쪽 oleic 산의 사슬은 palmitoleic 산성 (Δ 11 C16 ∶ 1) 그리고 oleic 산은 (Δ 13 C18 ∶ 1)다.일반적인 ω 7 지방산 (palmitoleic 산 (9 Δ C16 ∶ 1) 그리고 oleic 산은 (Δ 11 C18 ∶ 1))에서 많은 야생 식물과 조류에서 형성 되 할 수 있는 palmitoyl-ACP (C16 ∶ 0 ACP)에 의해 desaturation, polyketone과 다른 반응이다.슐츠 et al. [41] 다는 것을 발견 한 소설 유형의 geranyl ACP desaturase에 제라늄 dehydrogenizes 능 연구 acid-ACP (C14:0 ACP) 능 연구를 형성 하기 위해 acid-ACP (9 Δ C14:1), 그리고 두를 형성 하기 위해 polyketide과 다른 반응을 일 으 킨 불포화 지방산 (palmitoleic 산 (Δ 11 C16:1)와 oleic 산 (Δ 13 C18:1))다.따라서 은행골산의 전구체 지방산의 합성 경로는 제라늄의 펠라르고놀산의 합성 경로보다 탐색이 쉽다.
신갈 [38]은 제라늄의 단일불포화지방산 (팔미톨레산, 올레산) 합성과 관련된 유전자의 발현을 확인하고 확인하였다.그 결과 조직 내 acyl carrier protein (ACPs), ketoacyl synthases(KASs) 및 thioesterases (TEs)의 유전자 발현이 높을 때, 지방산과 ursolic acid의 함량도 높았다.Acyl carrier protein (ACP)은 지방산 합성 과정의 중심이 되는 보존적 중간 운반체이다.AAD (acyl-ACP desaturase)와 함께 작용하여 아실 사슬을 탈포화시켜 포화 지방산과 불포화 지방산의 비율을 변화시킨다.또한 ketoacyl-ACP synthase (KAS)와 함께 속도 제한 효소로 작용하여 페놀산 합성 속도를 조절할 수 있습니다.
2.1.2 Phytylbenzene 고리 합성
Phytylbenzene 고리 합성은 고지방산으로부터 페놀산을 형성하는 핵심 단계입니다.합성 경로는 그림 4에 표시되어 있습니다.을 gammacerolic 산 (C15:1) 예 로서, 첫째, palmitoyl (9 Δ C16:1)에서 지방산 경로 로서 malonyl-CoA 기판을 사용하 며 아래의 행동을 Polyketide Syn-thase Ⅲ (PKS Ⅲ), 4 개의 탄소 원자에 추가 결국 acyl 두 단계로 응축 반응을 통해 한다;둘째, C3위치의 케톤기는 케톤환원효소 (pks-ketoacyl-coa reductase, KR)에 의해 환원되어 히드록시기를 형성하고, 이중결합은 물 한 분자를 제거하여 이중결합을 형성하는 탈수증기 (PKS-dehydratase, DH;셋째, 응축 및 중합 후 아실 말단에 2개의 탄소 원자가 더 첨가된다.이때, C2위치의 수소이온은 4,15-디엔 트리펩타이드 중간체 구조의 안정성을 유지하기 위해 C7위치의 케톤 그룹에 더 가깝게 이동하며;넷째, C1의 케톤기가 탈카복실레이트가 되지 않도록 하면서, C2의 수소이온이 C7의 케톤기와 순환하는 것을 촉매하여 알돌응축과 유사하게하고, 탈수효소 (PKS-dehydratase;다섯째, 에노닐환원효소 (PKS-enoyl reductase, ER)는 육각형 탄소고리에 케톤기의 이중결합 및 방향족 형성을 촉매한다.와 C1 카르복실기가 함께 벤조산 구조를 형성하고;마지막으로 단일 불포화 측쇄가 형성되어 우르솔산 (C15:1)을 생성한다 [37, 39].
Plant 유형IIIpolyketide synthase (PKS III)는 페놀성 지질 (alkylphenols, alkylresor-cyls, urushiols, alkylcatechols 등) [42]의 합성을 위한 속도 제한 효소이다.Chalcone synthase (CHS)와 stilbenesynthase (STS)는 가장 대표적인 두 개의 슈퍼패밀리이다.STS)는 가장 대표적인 두 개의 슈퍼패밀리 중 하나이다.이들은 C1 및 C6위치 (클레이젠 응축)와 C2 및 C7위치 (알돌 응축)에서 각각 아실 사슬의 응축 및 순환화를 촉매하여 페놀성 물질을 형성할 수 있다.그러나 우론산은 C2위치의 수소이온의 응축을 촉매하여 C7위치의 케톤기를 갖는 동시에 C1위치의 카르복실기를 유지시켜 벤조산을 형성할 수 있다.벤젠 고리 카르복실기를 유지하는이 폴리케타이드 순환 반응은 식물에서 상대적으로 드물다.따라서 polyketide synthase type III(PKS III), cyclase (PKS-Cyclase) 및 ketoacyl reductase (PKS-ketoacyl-COA reductase, KR)에 대한 추가적인 연구를 통해 은행산 합성의 기전 (mechanism)을 더욱 밝힐 수 있으며, 은행조직 내 페놀산의 함량을 물리적, 화학적 또는 생명공학적 방법을 병행하여 조절할 수 있다.
페놀산 합성을 위한 주요 효소 유전자 2.2
At present, the main reports on the research of key enzyme genes for phenolic acid synthesis come from urushiol, while there are few reports on ginkgolides다.페놀산 합성 경로는 지방산 합성과 방향족 고리 합성의 2 부분으로 나뉜다.이 중 acyl carrier protein (ACP), stearoyl desaturase (SAD), ketoacyl-ACP synthase (KASs), type III polyketide synthase (PKS III), cyclase (PKS-Cyclase)는 우루시올 합성의 속도와 함량비를 결정하는 핵심 효소이다.
2.2.1 아실 캐리어 단백질 (Acyl carrier protein, ACP)
Acyl carrier protein (ACP)은 운반단백질의 대가족에 속한다.혼합단백질로서 다양한 단백질-효소 복합체와 결합하여 아실 사슬을 한 효소 중심 부위에서 다른 부위로 옮길 수 있다.또한 stearoyl-ACP desaturase (SAD) 및 acyl-ACP hydrolase (AAH)의 보조 인자로 작용하며, 지방산 합성 (FAS) 및 폴리케타이드 합성 (PKS) 경로에서 중요한 역할을 합니다.
이맹준 등 [43]은 Arabidopsis thaliana, Clycine max, Oryza sativa, Zea mays 등 17 종의 아실 운반체 단백질 유전자의 구조를 분석하여 5가지로 분류하였다.그 중 플라스미드형 ACP 유전자군 (코딩 영역은 4개의 엑손과 3개의 인트론으로 구성)과 미토콘드리아형 ACP 유전자군 (코딩 영역은 2개의 엑손과 1개의 인트론으로 구성)이 전체의 가장 큰 비중을 차지한다.그들은 둘다 4&와 결합할 수 있는 매우 보존적인 세린 사이트를 가지고 있다#39;-phosphopantetheine adduct cofactor group을 투여하고, holo-ACP를 활성화시켜 기능하게 한다.가족의 3차 구조 ACP 단백질은 대부분 일관성 있고 보존 되 4로 이루어 져 있 α helices다.3의 α helices (I, II, IV)은 서로 평행하게, 나선 α (III)을 수직하는 동안을 나선 센터에 충치 소수 성 구조를 형성하 제공하는 다양 한 acyl에 대한 보호 소수 성 구조 체인 [44]다.
α에 나선 Ⅱ ketoacyl-ACP과 교류 할 수 있는 synthase Ⅱ (β-Ketoacyl-ACP synthase Ⅱ, KAS Ⅱ) acyl 사슬을 연장 하기로 더, 그리고은 인정을 나선으로 알 려 져 있다.신갈 [38]은 제라늄에 대해 전사체 염기서열 분석을 수행하고 우루시올 산 합성과 관련된 두 개의 완전한 ACP 단백질 cDNA 서열 (Pxh1, Pxh2)을 걸러냈다.유전자 발현 검증 및 계통 발생 분석 결과 가 표명 된 Pxh1과 Pxh2 유전자들이 매우 제라늄의 trichome 조직에서, 그리고 높은 상동 14-carbon-1-ene에 산성 (9 Δ C14:1) biosynthesis ACP 단백질 유전자의 고수 (Coriandrum 알리 움), 어느 catalyzes 지방산의 desaturation uronic 산 경로에 있다.
2.2.2 Ketoacyl-ACP 합성효소 (KASs)
지방산의 아래에 조치는 synthase (FAS) 복잡 한, malonyl-ACP 기판의 전신을 형태로 사용 되는 ginkgolic 산성, palmitoleic 산성 (9 Δ C16:1)와 oleic 산성 (Δ 11 C18:1), 여러 응축 주기를 통해 합니다.지방산 합성효소 복합체는 cond-효소의 상과에 속하는 3개의 ketoacyl-ACP 합성효소 (KASs)로 구성되어 있다.이들은 모두 KAS 보존적 구조 도메인을 포함하는 소수성 지단백질이며 신호 펩타이드는 없다.KAS III는 말로닐-coa와 아세틸-coa의 중합을 촉매하여 초기 아실 사슬 (3-ketobutyryl-ACP;KAS I은 아실사슬과 말로닐-acp의 중합을 촉매하여 6C-16C로 지방산을 형성하며;그리고 KAS II는 malonyl-ACP와 palmitic acid의 응축을 촉매하여 18C 지방산을 형성합니다.

깻잎 [45], 고지대 [46] 등의 식물에서 카스 II는 아미노산 길이가 약 500 aa인 약산성 비분비 단백질이다.그것은 C16:C18 지방산의 비율을 결정하고 식물 내한성을 조절할 수 있습니다.연구원들은 7 얻 ketoacyl-ACP synthase (카스)의 transcriptome에서 유전자 Pelargonium xanthium, 그중 PxKAS Ⅰ 한, PxKAS Ⅰ b와 PxKAS Ⅰ c의 trichome 조직에서 높은고 안정 된 표현을 Pelargonium xanthium, 유전자 발현하는 동안에 다른 조직은 낮다.추가 치료의 trichomes 온도와라 디 언 트 (18, 23과 28 ° C)들의 표현의 다는 것을 보여주는 온도 가 증가와 감소 3 유전자,었고 긍정적으로 연관 되어 지방산의 내용 (palmitoleic 산 (Δ 11 C16:1)와 oleic 산 (Δ 13 C18:1))과 우 루시 올, 또는 우 루시 올 합성 경로에 참여하고 있다.진화론적 분석 결과, 제라늄의 KAS 유전자군은 Arabidopsis thailana 및 Glycine Max의 ketoacyl synthase (KAS) 유전자와 매우 상동성이 높고, 이와 관련된 촉매 메커니즘도 비교적 명확하다.그러나 ginkgolide ketoacyl-ACP synthase (KAS)에 대한 분자학적 연구는 거의 없었다.어떻게의 전조 palmitoleic 산성 (9 Δ C16:1)와 oleic 은행나무 산의 산성 (Δ 11 C18:1) (C15:1)와 heptadecenoic 은행나무 산성 (C17:1)은 규제를 받 ketoacyl-ACP synthase 가족으로 중요 monoenriched 불포화지방산을 깊이 탐구하는 것으로 남아 있다.
2.2.3 Stearoyl-ACP desaturase (SAD)
Stearoyl-ACP desaturase (SAD)는 식물에서 가용성 desaturase의 유일한 계열로 알려져 있으며, 포화지방산과 불포화지방산의 비율을 조절한다.그들 중, Δ 9 stearoyl-ACP desaturase은 가장 널리 식물에서 공부 했다.아실 사슬의 C9 및 C10위치의 탈수소를 촉매하여 첫 번째 이중 결합을 형성한다.SAD 단백질은 acyl-ACP desaturase 계열에 속하는 보존 도메인과 ferritin 계열에 속하는 보존 도메인으로 구성된 호모다이머이다.슬 픈 보존 되 지역에 4 α helices four-helix 번들을 형성 한 구조, 대칭을 숨 깁니다하는 Fe-O-Fe two-iron 클러스터 촉매 센터.이들은 함께 아실 사슬 탈수소 [47]를 위한 효소의 활성 중심을 형성한다.단백질 결정구조를 보면 SAD 효소는 분자 표면으로부터 내부까지 깊은 홈이 확장되어 있으며,이 홈은 18C acyl chain을 수용하고 홈 하단의 ferrous center와 결합하여 산화환원 반응을 할 수 있다.이 그루브 구조는 16C 및 14C 아실 사슬을 수용하여 반응할 수도 있지만 촉매 회전율이 낮다 [48].
Schultz 등 [41]은 제라늄 cDNA 라이브러리로부터 새로운 acyl-ACP desaturase를 스크리닝 하였다.유전자 염기서열 정렬 결과 피마자콩 stearoyl-ACP desaturase 유전자 (SAD)와 높은 상동성을 나타내었다.유전 변환 및 검증의 결과를 사용 하여 대장균 (대장균) 소설 유전자를 촉진 할 수 있 다는 것을 보여주 능 연구의 dehydrogenation acid-ACP (C14:0) 능 연구를 형성 하기 위해 acid-ACP (9 Δ C14:1), 그래서 그것은 능 연구 라는 이름의 acid-ACP desaturase 유전자 (9 화, Δ 14:0-ACP desaturase)다.Singhal [38] 유전자 발현 탐지 및 담배 (Nicotiana tabacum) 유전적 변환 유효성 검사 결과에 따르면 tetradecyl-ACP (9 Δ C14:1), 능 연구에 의해 촉발시 킨 제품 acid-ACP desaturase (MAD),은의 중요 한 전조 palmitoleic-ACP (Δ 11 C16:1)와 oleic-ACP (13 Δ C18:1)이다.의 활동 myristoyl-ACP desaturase (MAD, Δ 14 ∶ 0-9 ACP desaturase)의 콘 텐 츠 palmitoyl-ACP 결정 (Δ 11 C16 ∶ 1)과 oleoyl-ACP (Δ 13 C18 ∶ 1) 탐지 하여 효소 활동, 지방산 콘 텐 츠 그리고 페놀 산은 콘 텐 츠, 2의 내용을 차례로 조절 monoenyl 쪽 체인, oleic 산성 (C22:1과 C24:1)이다.게다가, Singhal 온도를 설정 [38] 그라 데이션 (18, 23 일과 28 ° C)과 관련 된 표현의 변화를 조사하는 효소 유전자 제라늄 trichome 조직의 수치를하고 있 다는 것을 발견 표현 능 연구 acid-ACP desaturase 유전자 (9 화, Δ 14 ∶ 0- ACP desaturase)와 stearoyl-ACP desaturase 유전자 (9 슬프다, Δ 18 ∶ 0-ACP desaturase) 발현은 온도가 증가함에 따라 감소하였다.
Wang et al. [49] cloned a stearoyl-ACP desaturase gene (SAD, Δ9 18:0-ACP desaturase) from Ginkgo biloba leaf cDNA and subjected Ginkgo biloba leaves to temperature stress (4, 15 and 45 °C). The results showed that the gene 표현was high at low temperatures (4 °C) and room temperature (15 °C), while the gene expression at high temperatures (45 ℃) was several times lower than that of the control group. Subsequently, Liu Xinliang et al. [50] showed that the GbSAD gene encodes a peptide chain with a chain length of 412 aa and a molecular weight of 47 kDa. Cluster analysis showed a high degree of similarity with the stearoyl-ACP desaturase (SAD) amino acid sequences of other gymnosperms. Exogenous hormone experiments showed that the expression of the GbSAD gene was not 규제by abscisic acid (ABA), methyl jasmonate (MeJA) or ethylene (ETH), but salicylic acid (SA) activated gene expression, with the highest expression value being 9.7 times that of the control group, indicating that SA may be 관련 된in the regulation of the fatty acid synthesis pathway.
Pelargonium graveolens 그리고 은행나무 포함 된 독특 한 능 연구 acid-ACP desaturase (9 화, Δ 14 ∶ 0-ACP desaturase)과 stearic acid-ACP desaturase (9 GbSAD, Δ 18 ∶ 0-ACP desaturase) 했다.두 효소 모두 온도에 민감하고 낮은 온도에서 높은 활성을 보인다.두 효소는 식물에서 유일한 수용성 탈포화효소이며 기능적인 구조 면에서 높은 유사성을 가지고 있다.그러므로, 상동 은행나무의 기능이 정상 검증 stearoyl-ACP desaturase (9 GbSAD, Δ 18:0-ACP desaturase) 추가 ginkgolide 산 합성의 메커니즘을 명확히 할 것이다.

2.2.4타입 III 폴리케타이드 합성효소 (PKS III)
폴리케타이드 synthases는 세 가지 범주로 나뉜다:모듈러 PKS로 알려진 ㉮ PKS I은 여러 가지 다기능 폴리펩티드로 구성되어 있으며, 이들 각각은 독특하고 재사용 불가능한 촉매 도메인을 지니고 있다.PKS II는 반복 또는 방향족 PKS 라고도 하며, 반복되는 반응 단계 동안 페놀 폴리케톤 구조가 여러 번 형성되는 것을 촉매하기 위해 재사용 가능한 도메인 집합을 사용하는 다중 효소 복합체의 반복 시스템입니다.chalcone synthase-type enzyme으로 알려진 PKS III는 처음 두 종류의 PKS 효소 계열과는 완전히 다르다.상동종 이중기능성 단백질을 재사용할 수 있으며, ACP의 활성화와 그 활성부위 4&에 의존하지 않는다#39;-phosphopantetheine sulfide 이며, ACP를 통해 acyl-CoA 기질을 활성화하여 acyl-CoA와 직접 반응할 필요가 없습니다.PKS의 종류에 따라 구조적 기작은 다르지만, 이들은 모두 C-C 결합의 형성을 촉매하기 위해 케토신타제 (ketosynthase, KS) 도메인 또는 서브유닛 (subunit)을 이용하여 디카복실레이트 (decarboxylate)와 아실릴-코아 (acyl-CoA)를 응축하여 탄소 사슬을 연장한다.
PKS III 계열은 매우 다양하다.Chalcone synthase (CHS)와 stilbene synthase (STS)는 가장 일찍 발견되고 가장 대표적인 두 계열이다.이들의 아미노산 서열은 60%~75% 가 비슷하다 [42].chalcone synthase (CHS) 계열은 식물에서 널리 발견된다.분자량이 40~45 kDa인 호모다이머 단백질이다.보존성이 높은 활성중심 (Cys-His-Asn의 조합)을 이용하여 3개의 피루브산-코아 기질을 이용하여 쿠마린 코아의 탄소 사슬 신장을 촉매하여 테트라펩타이드 중간고리 구조를 형성한다 [42, 51-52].테트라펩타이드 중간고리의 C6 수소이온은 C1 케톤기와 클레이센 응축 반응을 거쳐 탄소 헥사-멤버네이트된 고리를 형성하고, 이를 방향화하여 찰콘이라고도 알려진 디페닐케톤을 형성합니다 (그림 5). 찰콘은 플라보노이드 합성의 중요한 전구체입니다.stilbene synthase (STS) 효소는 식물과 미생물 (예:스트렙토마이세스, 효모 및 박테리아)에서 보고되었다.약 43 kDa의 분자량을 가지며, 2개의 소단위로 이루어진 이량체이며, chalcone synthase (CHS)와 상동성이 높은 특정 보존 도메인 IPNS(F) AGAIAGN을 갖는 단백질을 가지고 있다 [53].
STS 효소는 쿠마로일 코아를 기질로 사용하여 테트라펩타이드 중간고리 구조를 형성한다.테트라펩타이드 중간고리의 C7 케톤 그룹은 C2 수소이온과 알돌 응집반응을 거쳐 탄소 6원자 고리를 형성하고, 방향화를 거쳐 phytoalexin resveratrol glycoside [54]를 형성한다 (그림 5). 신갈 [38]은 제라늄 (Pelargonium hortorum)에서 제2 형 케토아실 코아 합성효소 (KCS2, Keto-acyl CoA synthase 2)를 확인하였는데, 그 3차 공간구조는 폴리케타이드 합성효소 제3 형 (PKS III)과 유사하며,그리고 우루시올 합성 경로에서 순환화 응축 반응에 관여하는 것으로 추측된다.Ketoacyl CoA synthase (KCS)는 매우 긴 사슬 지방산 (VLCFAs)의 합성에서 첫 번째 응축 반응을 촉매하는 속도 제한 효소입니다.유전자군에 대한 연구는 주로 모델식물인 Arabidopsis thaliana에 집중되어 왔으며, 다른 식물에서의 연구 보고는 거의 없다.Costaglioli 등 [55]은 Arabidopsis의 21개 KCS 유전자원을 유전자 상동성과 유전적 진화 분석을 바탕으로 4개의 subgroup (FAE1, KCS1, FDH, CER6)으로 분류하였다.그 중 FAE1 유전자는 James 등이 [56] transposon tagging 방법을 이용하여 복제를 한 최초의 KCS 계열 유전자였다.FAE1의 아미노산 서열은 다른 polyketide synthase (chalcone synthase (CHS), squalene synthase (STS) 및 type III ketoacyl-ACP synthase (KAS III))와 매우 상동성을 보인다.
종별로 Ketol-acyl CoA synthase (KCS)는 기질 특이성이 다르다.Ginkgolide A는에 속한다 옻칠 산성 물질 (lacquer acid substances 그리고 pathway specific type III polyketide synthase (PKS III)는 geranium&와 기능적 구조 면에서 유사하다#39;s type 2 keto-acyl CoA synthase 2). 또한 우루시올 산 경로의 polyketide synthase와 acyltransferase (STS)는 모두 tetrapeptide 중간고리에서 C7 케톤 그룹을 인식하고 C2 수소이온을 인식하여 알돌 응축 반응을 일으켜 탄소 hexa-membered ring을 형성한다.다른 점은 아실트랜스페라제 (STS) 가 알돌 응축 순환화 반응시 탈카르복실화 반응을 일으켜 C1위치에서 케톤기를 산화시켜 이산화탄소를 방출하지만, 락톤산의 폴리케타이드 순환화 반응시에는 C1 케톤기는 그대로 남아 있고, CoA 가 수소이온 (H+)으로 치환되어 벤조산 구조가 형성된다는 것이다.현재의 촉매 메커니즘은 아직 명확하지 않아 추가적인 연구가 필요하다.
2.2.5 폴리케타이드 사이클레이스 (PKS-cyclase)
2,4-dihydroxy-6-pentylbenzoic acid (Olivetolic acid) 라고도 알려져 있으며, 카나비노이드의 합성을위한 중요한 전구체이다.Gagne 등 [57]은 hexanoyl-CoA 가 대마초의 PKS III (TKS)에 의해 촉매되어 12개의 탄소를 포함하는 tetrapeptide 중간고리 구조를 합성하는 것을 발견하였다.TKS 효소가 촉매를 계속하면 C2 및 C7위치에서 아실 사슬이 응축되고 순환되면서 CO2를 방출하여 3,5-디 히드 록시-펜틸 벤젠 (올리브 톨)을 형성합니다;PKS cyclase (Olivetolic acid cy-classe, OAC)에 의해 촉매되는 응축 및 순환 반응이 2,4-디 히드 록시-6-펜틸 벤조산 (Olivetolic acid)을 수득하기 위해 C1 카르복실 그룹을 유지하는 응축 반응을 촉매하는 동안.단백질 기능 분석 OAC 효소에는 독특 한 다는 것을 보여 준 β-α-β-β-α-α-β acyl 그룹의 토폴로지 및 cyclization을 촉진 할 수 있 합 C2 및니다 위치에 카르 그룹 및 유지 합니다.
OAC 한 것은 새로 운 기능을 포함하는 단백질 dimeric α + β 배럴당 도메인 (dimeric α + β 배럴당 단백질, DABB),에서 발견 되는 주로인 박테리아, 곰팡이와 식물이다.대마초sativa의 cannabidiolic acid cyclase CsOAC을 제외하고, 식물 다브 유사 단백질들은 주로 heat stable protein (HS) 계열이며 스트롭토마이세스 종의 폴리케타이드 cyclase와 구조가 매우 유사하다 [58]..DABB 단백질은 작은 단백질 (12 kDa, 101 aa)로 PKS III와 함께 세포질에 위치하며, tetrapeptide 중간생성물의 접힘을 인도함으로써 보호자 같은 역할을 하며, 궁극적으로 벤조산 구조를 포함하는 물질을 형성한다.Gagne 등 [51]의 연구는 젖산 합성의 폴리케타이드 순환화를 위한 C1 케톤 그룹을 유지하여 벤조산 형성의 촉매 공정에 대한 새로운 참조와 아이디어를 제공했다.
3 전망
1960년대에 은행골라이드는 은행나무의 외부 종자 외피에 있는 활성 물질 중 하나로 밝혀졌다.고성능 액체크로마토그래피-질량분석기 (HPLC-MS/MS) 기술의 보급과 NMR(핵자기공명기술)의 검출 정확도 향상으로 은행잎, 열매와 같은 조직으로부터 5개의 일반적인 은행산과 4개의 은행골류를 분리, 식별할 수 있게 되었다.은행나무는 옻나무이며 독특한 극성 (소수성, 친수성)을 가지고 있어 의약, 화학, 미용, 해충 방제 등 여러 분야에서 사용되고 있다.특히 은행산 (C15:1)은 후쿠다 등에 의해 [59] 작은 유비쿼터스 관련 수정자 단백질 (SUMO)의 아실화를 막아 관련 세포 기능을 조절하는 것으로 나타났다.암 및 신경질환 치료의 유력한 약물로 평가되고 있어 연구 개발 가치가 있다.
우루시올산은 은행나무 외에도 캐슈나무 (C. equatorialis), 옻나무, 피스타치오너트, 제라늄 등의 상용작물에서도 분리 및 확인되었다.슐츠 등 [60]은 우루시올 산 합성에 관한 실험을 하였다.그 결과 14c로 표기된 구연산, propionyl-CoA, oleoyl-CoA, acetic acid, myristic acid 및 palmitic acid를 Pelargonium graveolens의 trichome 세포와 배양하였다.oleoyl-CoA (C18:1) 만이의 합성에 관여하는 것으로 밝혀졌다 효과적인 탄소원으로 urushiol을, 다른 물질은 triacylglycerol을 합성하여 저장지질을 형성하는 경향을 보였다.따라서 palmitoleoyl-CoA (C16:1)와 oleoyl-CoA (C18:1)는 라코산산 합성 연구에 중요한 전구물질로 확인되었다.그러나 ginkgolide는 은행나무 특유의 phytoalexin 으로서 특수하고 보존적인 분자 진화 합성 경로와 조절을 가지고 있다.현재 은행골리드 합성조절에 대한 연구는 주로 지질전사체 및 대사체 분석에 초점을 두고 있는 반면, 폴리케타이드 합성기작에 대한 탐구와 관련 유전자의 클로닝에 대한 보고는 거의 없어 추가적인 연구가 필요하다.
MYB와 WRKY 전사인자는 type III polyketide synthase (PKS III) 계열 성원의 발현을 조절하여 저항성 효과와 함께 이차대사산물의 함량 변화에 영향을 줄 수 있다.옻칠산 합성에 있어 핵심적인 율제한 효소는 type III polyketide synthase (PKS III)로 ketoacyl CoA synthase (KCS)와 상동성이 매우 높다.Eckermann 등 [61]은 제초제 metazachlor 가 polyketide synthase 계열 III의 chalcone synthase (CHS)와 squalene synthase (STS) 뿐만 아니라 very long chain fatty acid (VLFAS)의 합성에 있어서 속도제한효소 (ketolacton CoA synthase (KCS))를 불활성화시킬 수 있음을 발견하였다.클로로아세트아미드 메타자클로는 관련 효소의 주요부위에 있는 시스테인과 결합할수 있어 응결반응이 정상적으로 진행되지 못하게 할수 있다.앞으로 laconic acid를 합성하는 type III polyketide synthase (PKS III)도 불활성화 시킬 수 있는지 은행나무 정지 세포주를 이용하여 화학물질을 검증할 예정이다.이를 통해 젖산 합성에 있어서 type III polyketide synthase (PKS III)의 촉매 기능과 조절 메커니즘에 대한 추가적인 통찰을 제공할 것이다.이러한 조절기전의 점진적인 해명을 통해 페놀산의 생산량이 적거나 많은 세포주를 배양할 수 있는 이론적, 실무적 지침을 제공할 것이다.
참조
[1] 마징, 후오샤오치안, 천치안 등이 있다.Mpro와 PLP [J]를 기반으로 한 잠재적 항신종 코로나바이러스 한약 선별 연구.한의학회지 2020, 45(6):1219-1224.
[2] 왕수안, 강안, 디류칭 등이 있다.은행나무 빌로바 추출물의 주요 활성 성분의 약리 연구 진행 [J.한방의학, 2013, 44 (5):626-631.
[3] 장신희, 구혜선, 왕기빈 등.부제:Research progress of ginkgo biloba seed coat [J.헤이룽장 농학 2018(11):156-160.
[4] 이토 카 HTOTSUKA, N, NAKAHARA K, et al.Antitumor 은행나무 (은행나무) l.다 [J다]화학 & 제약 회보,1987,35 (7):3016-3020.
[5]GELLERMAN JL,SCHLENKH.은행나무 (Ginkgo biloba[J]) 로부터 14개의 c 표지지방산과 아나카르디산의 제조 (Preparation of 14 c 표지지방산 및 아나카르디산)Lipids,1969,4 (6):484-487.
[6] 겔러먼 JL, 앤더슨 W H, 쉬렌크 H.Ginkgo biloba[J]. lipids,1974,9(9):722-725 아세테이트로부터 아나카르딕산의 생합성 (Biosynthesis of anacardicacid from acetate in Ginkgo biloba).
[7] 겔러먼 J L, 앤더슨 W H, 쉬렌크 H.anacardic의 합성 Ginkgo biloba[J]의 씨앗에 있는 산.Biochimica et biophysica acta,1976, 431 (1):16-21.
[8] 왕 J, 유 B, 류 X 외.부제:Isolation and identification of the chemical components in the outer seed coat of Ginkgo biloba [J].한약학, 1995, 26(6):290-292, 328.
[9] 이홍청, 허자오판, 장영민 외.연구논문:은행나무 (Ginkgo biloba)의 외피 종자의 hydroxyphenolic 및 hydroxyphenolic acid 성분에 관한 연구 (J.한약학, 2004, 35(1):18-20.
[10] 코주벡 A, 타이먼 J H.생리활성 페놀성 지질 [J.Studies in natu-ral products chemistry,2005,30:111-190.
[11] 조조 Fuliang다.중국은행학회지 (The Ginkgo Biloba Journal of China [M.북경:중국림업출판사, 2007.
[12] [12] 헤스크 D, 크레이그 R, 뭄마 R O.간의 anacardic acid biosyn-마취능 비교 충성이 and-susceptible 제라늄다 [J다] 한국화학생생태학회지 1992,18(8):1349-1364.
[13] GEDAMP H, SAMPATHKUMARAN P S.캐 슈 너트 쉘 리퀴드:Ex-견인, 화학 and 응용 프로그램 [J].진행 in 유기 농 코팅, 1986,14 (2):115-157.
[14] 우하이하, 우케이, 류진다 등.Ginkgo biloba 씨앗으로부터 ginkgolides의 정제, 동정 및 항균활성 (Purification, identification and antibacterial activity of Ginkgo biloba seeds)한국식품과학회지 2015, 15(3):207-215.
[15] Tian ZP, Yu W, He GZ 등.HPLC에 의한 은행잎의 총은행산 측정 (Determination of total ginkgo acid in ginkgo biloba leaves by HPLC [J])미량원소와 건강연구, 2015, 32(2):36-37, 47.
[16] 그는 징그렌.은행산의 알레르기성 및 작용기전 연구.우한:화중농업대학, 2003.
[17] 류준풍.연구논문:은행가루로부터 은행골라이드를 제거하는 과정에 관한 연구 (a study on the process of removal of ginkgolides from ginkgo powder)Tai' 안:산동농업대학, 2017.
첸 [18]. H Y, 쳉 K C, 슈 R novel electrospun nanofiber mat에 고정화된 laccase에 의한 은행산의 효소분해 (J.한국식품영양과학회지 2020년,100(6):2705~2712.
[19] 리앙 리싱.은행나무 (金融樹)의 외피 (外方) 연구 현황 및 발전 전망중국자원종합이용, 2003, 21 (10):12-14.
자오청린 (趙成林) [20]은행나무 외투의 산성 성분의 추출과 약용적 고찰.한방의학, 1997, 28(4):250-251.
[21] 지율.은행나무 (ginkgo biloba) 씨코트 추출물에 의한 작물 병원성 세균 저해에 관한 기초 연구 (J.Anhui Agricultural Science, 2005, 33(9):1598-1603.
[22] 수이천, 통근, 구희룡 외.은행나무 외피종자에서 추출한 중간체에 의한 곰팡이 성장 억제에 관한 실험적 연구 (J.한의약, 1990, 13(6):36-37.
[23] 무로이 H, 쿠보 1세.anacardic acid와 totarol의 항균활성, a-lone and in 조합 with 에 대한 methicillin, 메 티 실린 황색 포도 상구 균다 [J다]저널 of 적용 bacteriology, 1996년, 80 (4): 387-394다.
[24] 쉬키티안.ginkgolides에 의한 농업해충의 방제 및 방제에 관한 연구.산림화학공업, 2004, 24 (2):84-87.
[25] 등예청, 서한홍, 레이링.3가지 농업해충에 대한 ginkgo biloba 추출물의 접촉독성 (J.남중농업대학 논문집 2004, 25(3):61-63.
[26] 힐 G A, 마타코티 V, 그레이엄 W D.옻나무 [J]의 독성원리.미국 화학 저널 (Journal of the American chemical 사회,1934,56(12): 2736-2738다.
[27] 청량, 루펑창.부제:Research overview of Ginkgo biloba [J].약학진보, 2004, 28(5):209-213.
[28] 빈치에리 F F, 빈첸치니 MT, 반니 P.Ginkgo bilobaseed의 sarcotesta에서 활성 com-파운드 추출:일부 dehydro-genases 활성 억제 [J].Journal article,2013,63(2):79-82.
[29]AHLEMEYER B,SELKE D,SCHAPER C,et al.Ginkgolic acid는 신경의 죽음을 유도한다 and 활성화 protein phosphatase type-2C다 [J다]유럽 약리학 저널,2001,430(1):1-7.
[30] 양장팅 (Yang Jianting), 우카이 (Wu Cae)은행나무 (Ginkgo biloba)의 알레르기성 성분과 감작기전에 관한 연구 진행 [J.식품공학, 2009, 34 (6):282-286.
[31]AL-YAHYA A A,AL-MAJED A A,AL-BEKAIRI A M, 외 스위스 알비노 생쥐 Ginkgo biloba의 생식, 세포학적 및 생화학적 독성에 관한 연구 (studies on the reproductive,cytological and biochemical toxicity of Ginkgo biloba in Swiss albino mice)한국민족약리학회지,2006,107 (2):222-228.
[32]JIANG L,SI Z H,LI M H,et al.1 H nmr 기반 대사체 연구 ginkgolicacid에 의해 유발된 간 손상에 대한 연구 (15 화~1 화) 쥐 [J.phar-maceutical and biomedical analysis 논문집,2017,136:44-54.
[33]야 오 Q Q,리 L, XU, M C, et al.The 의 대사 및 간 독성 ginkgolicacid(17 土 1 土) in vitro[J].중국 저널 자연의 의약품, 2018,16(11):829-837.
[34]QIAO L N,ZHENG J B,JIN X Z,et al.Ginkgolic acid는 AMPK 활성화를 통해 대장암세포의 inva-siveness를 억제한다.Oncology let-ters,2017,14 (5):5831-5838.
[35] LIANG J R, 양 H.Ginkgolic 산 (GA) 억제 위암 암 성장 by 유도 세포 사멸과 억압 에 의해 규제되는 STAT3/JAK2 신호 ROS[J/OL].Biomedicine & pharmacotherapy, 2020년, 125다 [2020-03-21다]https:∥ doi.org/10.1016/j.biopha.2019.109585 참조.
[36] 류이 X, 양 B, 장 L R 등 ginkgolic acid 가 인터플레이를 유도한다 apoptosis와 사이 autophagy regulated by ROS 세대 in 콜론 암 [J]이다.생화 학적 and 생물 물리학 research 커뮤니케이션,2018, 498(1):246-253.
[37] 월터스 D S, 크레이그 R, 뭄마 R O.제라늄의 아나카르딕산 생합성에 지방산 결합 [J.Phytochemistry,1990,29 (6):1815-1822.
[38] SINGHAL R A.식별 and 성격 묘사 유전 자의 involved in n5모노엔의 신진대사 선임자들 to n5 anacardic acids in the Pelargonium x hortorum[D]의 tri-chomes.루이빌:대학교 of 루이스-ville, 2016년.
[39]NARNOLIYA L K,KAUSHAL G,SINGH S P, 외. de novo transcriptome 분석의 장미 향기 geranium 신진대사에 대한 통찰력을 제공합니다 테르펜과 타르타르산 생합성의 특이성 [J.BMC Genomics,2017,18(1):1-14.
[40] Wu Yongmei, Mao Xue, Wang Shujian, 외.대사 공학의 식물 ω 7 지방산이다 [J다]「 Acta Botanica Sinica 」, 2011, 46(5):575-585.
[41] 슐츠 D J, CAHOON E B, 샨 크 J, et al.Expression Δ의 9 14: 0-acyl 캐리어 단백질 지방산 desaturase 필요 한 유전자이 프로-duction의 ω 5 해충저항성 제라늄 (pelargoni-um xhortorum) [J]에서 발견되는 아나카르산.절차 of the 医学 [명] 【 의학 】 医学 과학,1996,93(16):8771-8775.
[42]SHI S P,MORITA H,WANIBUCHI K,et. enzymatic synthesis of 식물polyketide [J].Current organic synthesis,2008,5(3):250-266.
[43] Li Mengjun, Shi Zhanliang, Guo Jinkao, 외.식물acyl carrier protein 유전자군의 염기서열 분석 [J.북방농업대학 논문집 2010, 25(S1):1-6.
[44]KOGLIN A,MOFID M R,LHR F, 외. conformational switches modu-late protein 상호작용을 in 펩타이드 항생제 합성효소 [J.과학, 2006,312 (5771):273-276.
[45] Li Lu, Liang Qian, An Xi 외.정보학 분석의 깻 β-ketoacyl ACP synthase 유전자 가족 [J]이다.산시 농학, 2017, 45(3):321-324.
[46] 하오칭팅.식별 및 기능 분석의 β-ketoacyl-ACP synthase II (KAS II) 가족은 고지대에 유전자 면화 [D]다.타이구 (泰區) 산시농업대학 (山西農業大學, Shanxi Agricultural University, 2018) 59명
[47]KACHROO A,SHANKLIN J,WHITTLE E,et al., the Arabidopsis stearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms 올레산 합성 [J]에.식물분자생물학,2007,63(2):257-271.
[48] 타하 R S, 이스마일 1세, 자날 Z 등 스테아로일-아실-캐리어-단백질 기름야자나무의 desaturase promoter(Des) 가 과실의 특이적 GUS expres-sion in transgenic tomato[J].식물생리학회지 2012년,169(13): 1290-1300다.
[49] 왕 H L, 조 F L, 장 X W, 외 al.Cloning and expression Ginkgo bi-loba L.[J]의 stearoyl-ACP desaturase와 두 oleate desaturases 유전자의.식물분자생물학기자,2013,31 (3):633-648.
[50] Liu Xinliang, Cai Jinfeng, Wang Huanli, 외.abiotic stress와 원핵생물 발현에 대한 Ginkgo biloba SAD 유전자의 반응 (J.동북임업대학 논문집 2015, 43(12):1-6.
[51] NAKANO C, 오자와 H, AKANUMA G, et al.Biosynthesis of Ba-cillus subtilis의 type III polyketide synthase와 methyltransferase에 의한 지방족 폴리케타이드 (J.한국세균학회지 2009,191 (15):4916-4923.
[52] JEZ J M, 오스틴 M B, 페러 J L, et, al.Structural control 식물 특이적 폴리케타이드 신테이스에서의 폴리케티 변형의 [J.화학 &생물학,2000,7 (12) :919-930.
[53] 류징잉, 둥쉐밍, 허우허성.qhe 유전자 (J.천진농학, 2015, 21(4):24-27.
[54]AUSTIN M B,BOWMAN M E,FERRER J, 외. an aldol switch discov-ered in stilbene synthases 중재 cyclization 특이성 of type III polyketide synthases다 [J다]화학생물학 (Chemistry & biology),2004,11 (9) :1179-1194.
[55]COSTAGLIOLI P,JOUBS J,GARCIA C,et. profiling 후보 유전자들 microarray analy-sis [J]에 의한 Arabidopsis thaliana에서 왁스 생합성에 관여한다.Biochimica et biophysica acta,2005,1734 (3):247-258.
[56]JAMES D W,JR,LIM E,KELLER J,et al.의 Arabi-dopsis FATTY ACID ELONGATION1(FAE1) 유전자의 maize transpo-son Activator (J)를 이용한 directed tagging.식물세포,1995,7 (3):309-319.
[57]GAGNE S J,STOUT J M,LIU E, 등의 olivetolicacid cyclase의 확인 Cannabis sativa 드러 낸 a unique 촉매 경로 to plant polyketides다 [J다]한국학술정보,2012,109 (31):12811-12816.
[58] 마이티, a.r., 그리고 마지.새로운 다기능 단백질 계열 DABB 연구 진행.Genomics and Applied Biology, 2018, 37(12):5460-5472.
[59] 후쿠다 1세, 이토 A, 히라이 G, et 알. 은행산은 단백질을 억제한다 E1-SUMO 중간체 [J]의 형성을 차단하여 su-moylation.화학-생물학 (chem-chemistry & biology),2009,16(2) :133-140.
[60] 슐츠 D J, 위크라마싱헤 N S, 클링게 C M.6장-아나카르딕 산 생합성 하고 [M] bioactivity ∥ 로미오 J T.최근 phytochemistry의 약자이다.암스테르담:Elsevier,2006:131-156.
[61]ECKERMANN C,MATTHES B,NIMTZ M,et alvalent binding of chlo-roacetamide 제초제 to the active 사이트 시 스 테인 of plant type III polyketide synthases다 [J다]Phytochemistry,2003,64 (6):1045-1054.
-
Prev
어떻게 Ginkgo Flavone을 추출하는가?
-
다음
Chamomile Extract Ingredient: Empowering Natural Cosmetics Formulation




