

 영어
영어 프랑스
프랑스 스페인
스페인 러시아
러시아 한국
한국 일본
일본Natural Beta Carotene Ingredient Elevates Animal Nutrition Formulas
today&에서#39; s pursuit of maximizing farming efficiency, how to transform every ounce of nutritional investment into tangible production outcomes is the foremost concern for every livestock manager다. 녹색 봄 Technology deeply understands your needs. Our natural beta-carotene solution, with its exceptional efficacy and scientifically optimized formulation, is emerging as a new choice for modern farms seeking to enhance profitability.
As the most efficient natural source of Vitamin A, beta-carotene holds irreplaceable value in enhancing animal health and improving production performance. However, traditional usage often faces challenges like low absorption efficiency and difficult dosage control, preventing this premium nutrient from reaching its full potential.
녹색 봄 Technology has overcome this challenge through innovative technology. Our uniquely formulated product significantly enhances beta-carotene absorption and utilization within animals, ensuring every gram delivers maximum value. Crucially, we provide precise dosage guidance tailored to different animal species, growth stages, and production goals, empowering you to effortlessly implement optimal usage protocols.
By choosing Green Spring Technology, you gain:
✓ Precise and efficient nutritional supplementation solutions
✓ Predictable improvements in production performance
✓ Professional technical guidance services
✓ Consistent product quality assurance
1 β-Carotene: Exceptional Structure and Properties Laying the Foundation for Precision Nutrition Science
today&에서#39;s rapidly advancing field of animal nutrition science, the molecular structure and properties of raw materials directly determine their nutritional efficacy. Leveraging in-depth analysis of the β-carotene molecular structure and its properties, Green Spring Technology has successfully overcome traditional application limitations through innovative technology, providing a robust scientific foundation for precision animal nutrition.
1.1 Unique Molecular Structure, Exceptional Nutritional Value
Green Spring Technology's premium natural β-carotene possesses distinctive molecular characteristics: a short straight-chain structure composed of 40 carbon atoms, featuring 15 conjugated double bonds and two β-violane rings at both ends. This unique configuration makes it nature's most potent and abundant vitamin A precursor. It efficiently converts into bioactive vitamin A within animals, providing critical nutritional support for animal health.
1.2Precise Classification for Optimal Efficacy
Based on carotenoid chemical properties, Green Spring Technology has established a scientific classification system:
· Carotenoid Class: Includes non-oxidized carotenoids such as β-carotene and α-carotene
· Lutein Class: Covers oxidized derivatives including zeaxanthin and astaxanthin
Our R&D team has confirmed through in-depth research that β-carotene exhibits the strongest bioactivity among numerous isomers, making it the optimal choice for animal nutritional supplementation.
1.3Innovative Technological Breakthroughs to Enhance Utilization Efficiency
Addressing the high hydrophobicity and nonpolar nature of β-carotene, Green Spring Technology has developed proprietary polarity optimization techniques:
· Enhancing hydrophilicity through molecular modification to improve absorption efficiency in animals
· Employing microemulsion technology to improve dispersibility, ensuring uniform distribution in feed
· Utilizing encapsulation protection technology to prevent activity loss during processing

1.4Scientifically Validated, Quality Assured
Green Spring Technology's natural β-carotene strictly adheres to scientific standards:
· Stable active ingredient content with less than 2% variation between batches
· Industry-leading vitamin A conversion efficiency
· Passed rigorous stability and bioavailability testing
2 Green Spring Technology Natural β-Carotene: A Full-Process Precision Nutrition System, Pioneering a New Era of Healthy Farming
In modern healthy farming systems, the absorption, transport, metabolism, and deposition efficiency of nutrients collectively determine final farming outcomes. Leveraging deep insights into the full metabolic pathway of β-carotene, Green Spring Technology has established a comprehensive precision nutrition solution, providing robust technical support for advancing scientific and precise animal nutrition.
2.1 Overcoming Absorption Bottlenecks to Establish Efficient Nutrition Foundations
Green Spring Technology has successfully broken through traditional absorption limitations with innovative technologies, laying a solid foundation for efficient animal nutrient utilization:
Precision Regulation of Absorption Efficiency
Leveraging the close correlation between β-carotene absorption and lipid content, we developed a proprietary lipid-enhanced formulation. By optimizing lipid ratios and utilizing micronization technology, we elevated β-carotene absorption efficiency to industry-leading levels. Our unique microemulsion technology ensures active ingredients maintain optimal bioavailability throughout the digestive tract.
Multi-species Adaptation Solutions
· Monogastric Animals: Optimized lipid ratios enhance intestinal absorption efficiency
· Ruminants: Specialized coating technology protects active ingredients during rumen transit
· Aquatic Animals: Micronization processes adapt to unique digestive physiology
2.2 Innovative Delivery Systems for Precise Targeted Transport
Green Spring Technology has established an efficient in vivo delivery system ensuring precise nutrient targeting to specific tissues:
Lipoprotein Precision Delivery
Based on species-specific physiological characteristics, we developed proprietary lipid complexation technology significantly enhancing β-carotene binding efficiency with diverse lipoproteins. By precisely regulating lipid composition, we ensure active ingredients efficiently enter key carriers, enabling effective transport from the gut to target tissues.
Species-Specific Transport Solutions
· Ruminants: Optimized high-density lipoprotein (HDL) delivery system
· Monogastric animals: Enhanced low-density lipoprotein (LDL) binding capacity
· Poultry: Balanced lipoprotein ratios for optimal transport efficacy

2.3 Precision Metabolic Regulation, Optimizing Nutrient Conversion Pathways
Green Spring Technology employs innovative techniques to precisely regulate metabolic processes, maximizing nutritional value:
High-Efficiency Vitamin A Conversion
Through in-depth research on key metabolic enzymes, we developed unique metabolic regulation technology that significantly enhances the conversion efficiency of β-carotene to vitamin A. By optimizing molecular structure, we ensure more effective recognition and conversion by metabolic enzymes.
Synergistic Multi-Physiological Functions
· Gene Expression Regulation: Metabolites participate in regulating target gene transcription
· Lipid Metabolism Modulation: Influences fat metabolism through specific signaling pathways
· Antioxidant Protection: Complete molecules provide sustained antioxidant support
2.4 Targeted Deposition Technology, Enhancing Tissue Nutrient Reserves
Green Spring Technology innovatively achieves directed deposition of nutrients in key tissues, significantly boosting nutritional efficacy:
Tissue-Specific Deposition
· Liver Nutrition Enhancement: Strengthens specific deposition in the liver
· Skin and Feather Health: Increases distribution concentration in skin tissue
· Reproductive Nutrition Support: Targeted enhancement of deposition efficiency in reproductive organs
Scientifically Validated Superior Efficacy
Extensive practical applications confirm animals using Green Spring Technology products demonstrate outstanding performance in:
· Significantly elevated liver vitamin A reserves
· Noticeably improved skin and feather coloration
· Comprehensive enhancement of reproductive system health
· Sustained optimization of overall production performance
2.5 Comprehensive Technical Service System
Green Spring Technology has established a robust technical support system, providing clients with:
· Customized Formulation Design: Precision solutions tailored to species and growth stages
· Production Process Optimization: Assistance in resolving technical challenges during manufacturing
· Effect Monitoring & Evaluation: Full-cycle technical tracking and service support
· Scientific Application Guidance: Ensuring optimal product performance
Green Spring Technology is committed to advancing healthy animal husbandry to higher levels through end-to-end precision nutrition solutions. We look forward to collaborating with industry partners to pioneer a new era of scientific and precision animal nutrition, contributing to sustainable development goals.
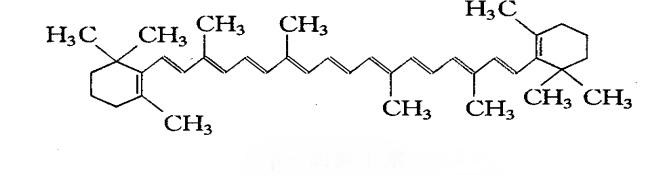
3 Green Spring Technology Natural β-Carotene: Multi-Dimensional Benefits Empowering Healthy Aquaculture
In modern healthy aquaculture systems, precision nutrition has become the core driver for enhancing farming efficiency. Green Spring Technology's Natural β-Carotene, systematically validated for its exceptional performance across three critical domains—production efficiency, immune regulation, and reproductive efficacy—delivers comprehensive nutritional solutions for farming enterprises, ushering the industry into a new era of scientific feeding.
3.1 Enhanced Production Performance: Data-Driven Efficiency Gains
Significant Improvement in Milk Production
· Supplementation of 400mg/day during heat stress increased milk yield by up to 11%
· Un데 르standard feeding conditions, 90mg/day supplementation increased milk yield by 11.03%-13.83%
· Continuous use maintains high and stable production, enhancing farm productivity stability
Comprehensive Dairy Quality Optimization
· Significantly increased milk fat content, improving dairy product flavor and nutritional value
· Milk protein content increased from 2.90% to 2.96%, enhancing market competitiveness
· Substantially increased milk solids content optimizes physicochemical product parameters
Precision Feeding Protocols
· Standard Enhancement Protocol: 90-300mg/day for balanced cost-effectiveness
· High-Yield Optimization Protocol: 400-900mg/day to maximize production potential
· Stage-Adjustment Protocol: Dynamic optimization based on lactation cycle for sustained returns
3.2 Immune System Fortification: Building a Health Defense System
Triple Immune Protection Barrier
· Enhanced Cellular Immunity: Promotes lymphocyte proliferation and strengthens immune response capacity
· Optimized Humoral Immunity: Increases immunoglobulin IgA and IgG concentrations
· Strengthened Non-Specific Immunity: Elevates serum lysozyme activity to establish broad-spectrum protection
Significant Disease Prevention Outcomes
· Substantially reduced incidence of retained placenta
· Effectively controlled risks of metritis and mastitis
· Reduced somatic cell count in milk by 18.54%-46.10%, enhancing dairy product safety and quality
Precision Immunization Protocol
· Dairy Herds: Prenatal supplementation boosts colostrum quality
· Poultry Farming: Promotes growth and development while enhancing overall immunity
· Critical Periods: Rapidly restores immune function to counter stress challenges
3.3 Reproductive Performance Optimization: Enhancing Herd Development Potential
Comprehensive Ovarian Function Improvement
· Increases luteal β-carotene concentration to promote progesterone secretion
· Peripartum supplementation of 500-2000mg/day significantly boosts ovulation rates
· Effectively prevents ovarian cysts; serum concentrations ≥33μg/dL significantly reduce herd incidence
Significantly improved embryo quality
· At serum concentrations exceeding 200 μg/dL, normal transferable embryos significantly increase
· Total recoverable embryos show stable growth
· Noticeably improved embryo quality boosts reproductive success rates
Scientific Reproductive Management
· Peripartum Protocol: Supplementation starting 3 weeks pre-partum optimizes ovulation outcomes
· Heat Stress Protocol: Continuous 400 mg/day supplementation enhances pregnancy rates
· Customized protocols: Tailored plans based on species-specific requirements
3.4 Demonstrated Comprehensive Benefits: Creating Sustained Farming Value
Green Spring Technology's natural β-carotene delivers holistic value enhancement for farming enterprises through the synergistic action of three core functions:
Significant Economic Benefits
· Enhanced production performance directly increases operating revenue
· Optimized reproductive efficiency reduces replacement costs
· Reduced disease incidence lowers medical expenditures
Outstanding Management Benefits
· Improved production stability facilitates planned management
· Enhanced herd health reduces unexpected incidents
· Optimized breeding cycles improve management efficiency
Sustainability Assurance
· Reduced antibiotic use ensures product safety
· Improved animal welfare aligns with modern farming principles
· Optimized resource utilization achieves efficient production
3.5 Professional Technical Support: Comprehensive Service Assurance
Green Spring Technology has established a robust technical service system:
· Customized formulation services
· Comprehensive technical guidance and monitoring
· Effectiveness tracking and solution optimization
· Professional technical training support
We understand that success in modern animal husbandry stems from precise control at every stage. Green Spring Technology is committed to providing partners with holistic support—from production to reproduction, and from immunity to management—through scientifically validated nutritional solutions. Choosing us means choosing to walk alongside science and achieve mutual success.

4 Green Spring Technology: Driving Scientific Advancements in Animal Nutrition Through Continuous Innovation
Scientific research has conclusively demonstrated that natural β-carotene is not only an efficient vitamin A precursor but also delivers multifaceted benefits in enhancing animal production performance, improving reproductive efficiency, and strengthening immune function. As the industry deepens its understanding of precision nutrition, the scientific application of β-carotene is emerging as a key technology propelling the development of healthy animal husbandry.
Green Spring Technology remains at the forefront of animal nutrition research. Leveraging continuous R&D investment and technological expertise, we are dedicated to providing more precise β-carotene nutrition solutions for diverse livestock species and production stages.
We understand that the path to scientific farming requires ongoing exploration. Moving forward, we will continue collaborating with industry partners to deepen β-carotene research in animal nutrition, jointly propelling the healthy farming industry toward a new era of greater efficiency and sustainability.
Take action now and embark on a new chapter in scientific farming!
We sincerely invite you to explore Green Spring Technology's natural β-carotene solutions:
Phone: +86 29 88313578
Mobile/WhatsApp: +86 13649243917
Email: helen@greenspringbio.com
Website: https://www.greenspringnatural.com/
Exclusive Technical Services:
✓ Free sample trials
✓ Customized formulation design
✓ Technical application guidance
✓ Full-process tracking services
Our professional technical team will provide one-on-one solutions to help you maximize aquaculture efficiency. Let's collaborate to empower the aquaculture industry with technology and build a brighter future together!
참조:
[1] Wang Shaoxia, Tang Xiaohua, Zhou Yonghong, 외.파프리카 적색의 추출공정에 관한 연구 (J.식품산업과학기술, 2008, 29(8):196-199.
[2] 본 린티그 J,SIES H.Carotenoids[J].Archives of Biochemistry and Biophysics,2013,539(2):99-101.
[3] 얀 Hongxiang다.의 소화와 흡수에 관 한 연구 β-carotene 위장 기관에의 염소와 항 산화 효과 [D]다.Master' s 논문.양주:양주대학교, 2007.
[4] 본 오일러 H, 카러 P, 크라우스 第 1 項 및 第 2 項의 規定에 관한 硏究 biochemie der tomatenfarbstoffe다 [J다]Helvetica Chimica Acta,1931,14(1):154-162.
-
Prev
Natural Beta-Carotene for Gel Candies Delivering Uniform Color Without Spotting
-
다음
Innovative Applications of Natural Beta-Carotene in Animal Nutrition Formulations




