

 영어
영어 프랑스
프랑스 스페인
스페인 러시아
러시아 한국
한국 일본
일본히알루론산이 멜라닌 대사를 조절한다는 연구
비정상적인 멜라닌 합성과 대사는 멜라스마와 다른 색소 질환 발생의 주요 기전이며, 광노화 피부의 중요한 특징 중 하나이기도 합니다 [1-2].경제발전과 피플&의 향상으로#39;s living standard, there is an increasing dem그리고을healthy 그리고youthful skin, 그리고how 을better regulate melan에서metabolism is 의great clinical significance. 루 론 산 has a wide range 의biomedical applications due to 의good tolerance 그리고non-immunogenicity. In recent years, it has been used as a filler 에서aesthetic medicine, playing an important role 에서facial rejuvenati에그리고improvement 의피부imperfections[3] . Currently, little research has been reported on the relationship between 루 론acid and melan에서metabolism. Whether hyaluronic acid can regulate melanin metabolism deserves in-depth study. In this paper, we will review the roles 의hyaluronic acid and cells involved in melanin biometabolism in recent years, so as to clarify the correlation between hyaluronic acid and melanin biometabolism, and to provide information for further research.
히알루론산의 생물학적 특성 1
Hyaluronic acid is a non-sulfated glycosaminoglycan composed of repeating disaccharide units of D-glucuronic acid and N-acetylglucosamine. It has a wide range of 분자weights, and its concentration and distribution in the 인간body vary according to tissue type, age, and disease status[4] . Hyaluronic acid acts as a lubricant and shock absorber in the skin and joints due to its viscosity, elasticity and other rheological properties[5] ; it also participates in the regulation of cellular 기능by activating intracellular 신호pathways 을 통해binding to hyaluronic acid receptors (e.g., CD44) on the cell surface. The size of the hyaluronic acid molecule is a major determinant of the activation of receptor-mediated signalling pathways, and only hyaluronic acid chains of a specific mass range are able to activate the corresponding receptor-mediated cell signalling, which is regulated by the balance between hyaluronic acid biosynthesis and degradation [4,6].
2 멜라닌 생체표층증은 다양한 요인에 의해 조절된다.
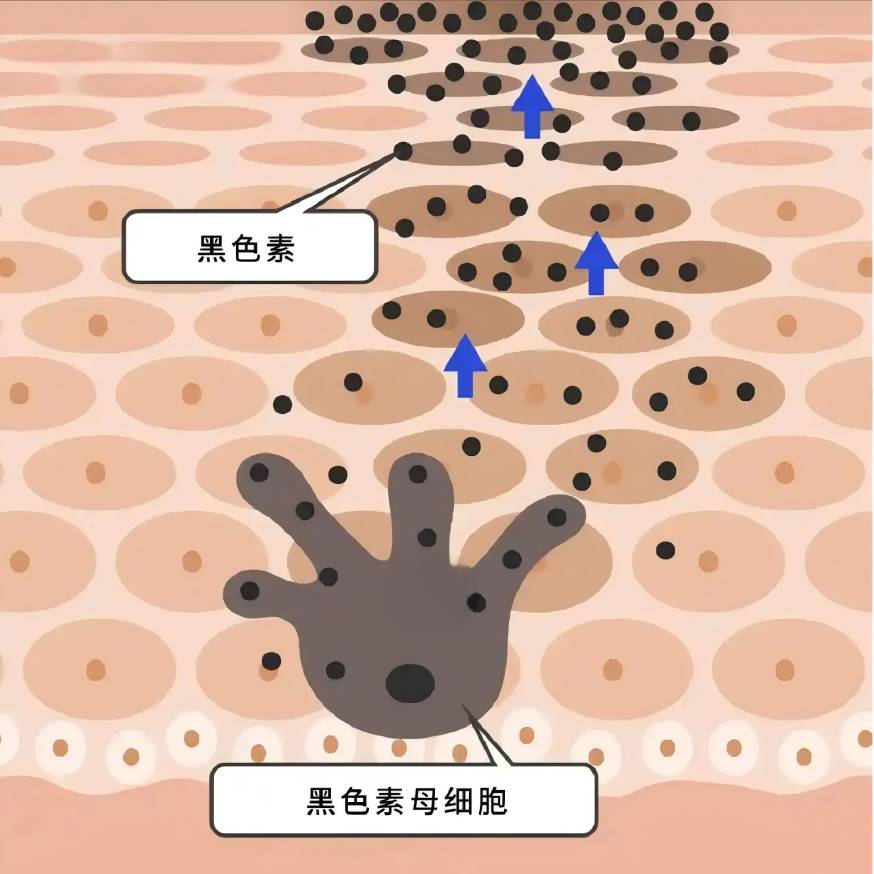
멜라닌은 멜라닌 세포에서 합성되어 각질세포로 운반되며, 다양한 세포내 및 세포외 인자들이 멜라닌 세포의 증식 및 생존 능력, 수상돌기 형성, 멜라닌 합성, 멜라닌소체의 수송 등에 영향을 미침으로써 긍정적 또는 부정적으로 멜라닌 생체 표상 조절에 관여한다 [7-8].
염증과 산화 스트레스는 멜라닌 합성에 영향을 미친다:염증 후 과색소침착에서 멜라닌 세포는 세포 증식과 활성을 증가시켜 염증에 반응하고, 멜라닌 생성이 증가하여 수상돌기를 통해 이웃한 각질형성세포로 옮겨져 과색소침착을 일으킨다 [9].;자외선은 세포막의 지질 과산화를 유발하며, 이는 멜라노사이트의 활성산소종 (reactive oxygen species, ROS)의 수준을 증가시켜 [10] 멜라노사이트의 과색소 형성을 자극할 수 있다.이는 멜라닌 세포를 자극하여 과도한 양의 멜라닌을 생성하게 할 수 있다 [10];백반증의 발병 원인에서, ROS는 분자 및 세포 기관 기능 장애를 유발하여 더 많은 면역 반응을 유발하고 궁극적으로 멜라노사이트 사멸 [11]을 초래한다는 것이 널리 받아들여지고 있다.
각질세포, 섬유모세포, 면역세포와 같은 멜라닌 세포에 인접한 세포들은 파라크린 인자의 분비를 통해 멜라닌의 대사 조절에 관여한다 [8,12-13].색소성 피부질환은 여러 멜라닌세포인접세포로부터 유래된 사이토카인의 비정상적인 발현을 동반하는 경우가 많다:흑점의 과색소 부위의 표피에서는 stem cell factor (SCF)와 각질형성세포로부터 유래된 endothelin-1의 발현이 up-regulated 된다 [14];멜라스마의 과색소 피부병변에서는 섬유모세포에서 추출한 분비성 프리즐드 연관 단백질 2 (sFRP2)의 발현 수준이 상당히 상향 조절된다 [15].(sFRP2)는 섬유모유 유래 분비물 프리즐드 관련 단백질로 상당히 상향조절된다 [15].전통적인 paracrine 요인 포함 keratinocyte-derivedα-MSH, 기본 섬유 아세 포 성장 인자 (FGF-2), fibroblast-derivedkeratinocyte성장 인자 (KGF), 분비 되는과 단백질 Dickkopf-1 (DKK1), 다른 것들을다 [13, 16-다]히스타민, 프로스타글란딘 E2, IL-6, IL-17 그리고 종양괴사인자 (tumour necrosis factor, TNF)와 같은 다양한 염증 매개체들도 멜라노 발생의 조절에 관여한다 [19-20].최근 멜라노사이트 이웃 세포의 파라크린 기능에 관여하는 인자들이 점점 더 많은 것으로 밝혀지고 있다 [21-23].
멜라닌 생균증의 히알루로난 조절 가능 기전 3
Lim et al[24] designed a self-controlled study using hyaluronic acid to fill the tear troughs. 4 weeks after topical 주사of hyaluronic acid, a decrease in localised melanin content as measured by a skin melanometer was observed compared to the control side, suggesting the potential of hyaluronic acid to influence melanin biometabolism.
3.1 히알루론산이 멜라노세포에 미치는 직접적인 효과
그것은 밝혀졌다히알루론산 0.2-5 g/L멜라노 세포의 증식을 촉진하고 멜라노 세포의 티로시나아제 활성을 증가시켜 멜라닌의 합성을 촉진 할 수 있습니다;10 g/L 히알루론산은 멜라닌 세포의 증식을 억제하고, 티로시나제 활성을 감소시키며, 멜라닌 합성을 억제할 수 있다 [25].
Hyaluronic acid has significant anti-inflammatory and antioxidant effects[26-27] : some studies have suggested that저분자량 히알루론산induces inflammation, while high molecular weight hyaluronic acid has an anti-inflammatory effect.26 Takabe et al.[28] found that UV treatment of melanocytes resulted in the inhibition of hyaluronic acid synthesis and the expression of the receptor CD44, and the inhibition of IL6, IL8, CXCL1, CXL1, and CXL1 수용체에melanocytes, IL6, IL8, CXCL1 그리고 CXCL10release in melanocytes; UV light and hyaluronanhydrolase co-treatment of melanocytes increased the release of the above inflammatory factors; exogenous addition of hyaluronic acid tablets did not significantly alter the 생산of inflammatory factors; the above results suggested that the unhydrolysed hyaluronic acid, which was present in the periphery of the melanocytes, could play a protective role in the inflammatory response of melanocytes, but the study did not focus on the role of the hydrolysis in this process in the melanocytic inflammation. The results suggest that hyaluronic acid, which is present around melanocytes and is not hydrolysed, could play a protective role in the inflammatory response of melanocytes, but the study did not focus on whether melanin synthesis was affected in this process. In the skin, hyaluronic acid acts as an antioxidant and free radical inhibitor[29]. .
In our previous study, we observed that hyaluronic acid intervention down-regulated ROS levels in 정상적인and aged fibroblasts, and Campo et al.[30] found that the addition of hyaluronic acid to the 산화system induced by Fe2+ and ascorbate in fibroblasts inhibited lipid peroxidation through chelation, suppressed ROS production, reduced DNA fragmentation and protein oxidation, and limited cellular growth through the addition of hyaluronic acid. The addition of hyaluronic acid to the ascorbate-inducedROS-generating oxidative system can inhibit lipid peroxidation through chelation, inhibit ROS production, reduce DNA fragmentation and protein oxidation, and limit cell death, thus exerting antioxidant effects. It is not known whether hyaluronic acid can directly exert an antioxidant effect on melanocytes in the current study, and further investigation is needed.
멜라노사이트 이웃세포의 파라크린 기능 조절에 히알루론산 관여 3.2
In previous studies, hyaluronic acid has attracted much attention because of its role in inflammation and wound healing[26,32] . Recently, it has been found that hyaluronic acid can regulate the expression and secretion of growth factors and interleukins by acting on keratinocytes, fibroblasts and immune cells in the skin.
3.2.1 히알루론산이 각질형성세포의 파라크린 작용에 미치는 효과
In the keratinocytes of aging mice, the binding of hyaluronic acid fragments (no more than 27 kDa) to its receptor CD44촉진RhoA-ROK signalling, while the interaction between high molecular hyaluronic acid (700 ~ 1 000 kDa) and CD44 자극하는activation of Rac-PKNγ, which affects the synthesis and secretion of keratinocytes [33]. UVB irradiation induces the degradation of hyaluronic acid into smaller fragments and selectively 활성화different cellular signalling mediated by CD44 in keratinocytes[34] .
In vitro studies have shown that 1% of high molecular hyaluronic acid (700-1,700 kDa) can increase the secretion of α-MSH from keratinocytes into the supernatant[35] . Hyaluronic acid oligosaccharides inhibited the release of IL-8 and TNF-α from LL-37-treated keratinocytes[36] . Hu et al.[37] found that very low molecular hyaluronic acid (0.8 kDa) and high molecular hyaluronic acid (1,200 kDa) inhibited the secretion of IL-6, IL-8, and IL-1 β by 인간표피keratinocytes after UVB irradiation, and the process involves the effect of hyaluronic acid on the secretion of TLRS and IL-1β in human epi피부keratinocytes[38] . This process involves the blockage of TLR4 activation by hyaluronic acid.
히알루론산이 섬유아세포 파라크린 작용에 미치는 효과 3.2.2
David-Raoudi et al.[38] investigated the 효과of natural hyaluronic acid, 12-and 880-saccharide hyaluronic acid fragments on the proliferation of human fibroblasts and the expression of matrix-related genes, and found that all three types of hyaluronic acid could promote the adhesion and proliferation of cells, and increase the expression of 행렬metalloproteinase-1 and matrix metalloproteinase-3, among which 12-saccharide hyaluronic acid could enhance the expression of type I collagen and TGFF-3 in fibroblasts. In particular, 12-saccharide hyaluronic acid enhances the expression of fibroblast type I collagen and TGF-β1.
Ciccone et al.39 reported that FGF-2 expression by fibroblasts was up-regulated when fibroblasts were exposed to 1 mg/mL of soluble hyaluronic acid, whereas the same concentration of a mixture of reticulated and free low-molecular-weight hyaluronic acid did not result in such an alteration.Asparuhova et al.[40] found that hyaluronic acid at 2,500 kDa (4 mg/mL) significantly increased the expression of human oral fibroblasts. Asparuhova et al.[40] found that 2,500 kDa of hyaluronic acid (4 mg/mL) significantly increased the expression of FGF-2, EGF2, IL-1 α, IL-1 β, and TNF in fibroblasts of human oral origin, and Quan et al.[41] showed an increase in the number of fibroblasts in the dermis, elongation of their morphology, and an increase in CCN2 in the dermis by subcutaneous injection of hyaluronic acid.
Radrezzaet al.[42] applied 양적 오 믹스to study the changes in protein profiles of normal human dermal fibroblasts with different concentrations of low molecular weight hyaluronic acid in the range of 20-50 kDa (0.125%, 0.25%, and 0.50%) and found that treatment with low molecular weight hyaluronic acid promoted the growth and proliferation of the cells, and increased the biosynthesis of proteoglycans, and the treatment of low molecular weight hyaluronic acid promoted cell growth and proliferation, and increased protein glycan biosynthesis at the highest concentration (0.50%). At the highest concentration (0.50%), inflammatory and immune responses were activated and the secretion of interleukins (IL-12, IL-1, IL-2, IL-4, etc.) and TNF-α was increased.
Lorén et al.[43] found that a 4.3-kDa hyaluronic acid fragment promoted the release of IL-6 and IL-8 from dermal fibroblasts 을 통해the CD44 receptor, but Olsson et al.[44] found that no increase in the release of IL-1 β, IL-6, IL-8, IL-10, IL-12, or TNF-α was detected after hyaluronic acid stimulation of fibroblasts from synovial membranes. IL-1β, IL-6, IL-8, IL-10, IL-12 or TNF-α were not detected after hyaluronic acid stimulation.
3. 2. 히알루론산이 면역세포의 파라크린 효과에 미치는 영향 3
면역세포에서 분비되는 염증인자도 멜라닌 합성 조절에 중요한 역할을 한다.그것은 스 루 론 산의 분비를 억제 할 수 없 다는 것을 발견, IL-6과 TNF-α 원시 264.7 LPS-induced 세포 내에;보조 T 세포에 사용될 때, low molecular weight hyaluronic acid 주로 IFN-γ의 분비를 억제하는 반면, 높은 분자 수준의 억제 루 론 산 무게를 IL-4 [45].
3. 2. 4 히알루론산은 이웃한 세포의 파라크린 작용을 조절함으로써 멜라닌 세포의 기능을 조절한다
On the basis of the previous theory, we designed a fibroblast-멜라 노 사이트co-culture system, and after photo-aging of fibroblasts from human skin, we found that the intervention of hyaluronic acid at 1 mg/mL on photo-aged fibroblasts inhibited melanin synthesis in melanocytes in the co-culture system, but did not have any significant effect on the melanin synthesis in the normal fibroblast-melanocyte system, which preliminarily proved that under certain conditions, hyaluronic acid inhibits the synthesis of melanin in fibroblasts and melanocytes. Under certain conditions, hyaluronic acid can regulate the 함수of melanocytes by regulating the paracrine action of fibroblasts.
조우 et al [46]의 UVB 방사선 조사 가 keratinocytes 유도 IL-18의 생산과 IFN-γ 복용 량-그리고 한 동 한 방법으로, 그리고 IL-18 멜라 노 사이트의 멜라닌 내용이 상당히 증가와 co-cultured keratinocytes, IFN을 때-γ 유의 한 억제 효과를 보였다.D'agostino et al [47] found that a mixture of hyaluronic acid and chondroitin promoted the increase of melanin synthesis in keratinocyte-melanocyte co-cultures, but chondroitin or hyaluronic acid alone did not have a significant effect on melanin content. This may suggest that hyaluronic acid needs to be in a certain form or under a certain condition in order to play its role in regulating melanin synthesis.
4 요약 및 전망
Although hyaluronic acid has been used for a long time and in a wide range of applications, its effects are not well understood because they are affected by a variety of factors such as molecular weight, cross-linking degree, concentration, receptor type, target cell type, etc. In the field of dermatology, hyaluronic acid has been used for a long time and in a wide range of applications. In the field of dermatology, few studies have been reported on the effects of hyaluronic acid on melanin biometabolism. The results of the existing clinical and basic studies suggest that hyaluronic acid may be related to melanin metabolism, especially the metabolic changes of melanin under external stimuli (e.g., UV irradiation), which is worthy of in-depth study. A deeper understanding of the functions of hyaluronic acid올바른 히알루론산의 유형을 더 잘 선택하고 미용의학과 색소성 질환 치료에 그 기능을 깨닫도록 도울 것입니다.

참조
[1] Passeron T, Picardo M.Melasma, 말하 장애 [J]다.돼지-ment Cell Melanoma Res,2018년,31 (4):461-465.
[2] 한아 (Han A), 치엔알 (Chien AL), 강 S.Photoaging[J.2014년 Dermatol Clin, 32 (3):291-299다.
[3] Bukhari SNA,RoswandI NL,Waqas M,et al. 히알루론산,a 유망한 피부 회춘 생의학:최근 업데이트 리뷰 그리고 화장품 및 뉴트리코스-메틱 효과에 대한 전 임상 및 임상 조사 [J].정수 J Biol Macromol, 2018년, 120 1682년 (광해군 2) 진사 (進士) 가 되었다 -1695.
[4] KobayashiT, ChanmeeT Itano N.Hyaluronan:신진대사와 기능 [J.생체 분자, 2020년, 1525년 10 (11):다.
[5]Cowman MK,Lee HG,Schwertfeger KL,et al.의 내용과 크기 히알루로난의 생물학적 수분 and 조직 [J]이다.앞 Immunol, 2015년, 6:261.
[6]Weigel PH.Planning,evaluating and vetting receptor 신호stud-ies to assess hyaluronansize-dependence and 특이성 [J].Glyco-biology,2017,27 (9):796-799.
[7] D'mello SA,Finlay GJ,Baguley BC,et al.Signaling pathways in melanogenesis[J]. 「 melanogenesis 」 (영어).Int J Mol Sci,2016,17 (7):1144.
[8]Pillaiyar T,Manickam M,Jung SH.Recent development of signaling pathways inhibitors of melanogenesis[J.「 Cell Signal 」,2017,40:99~115.
[9] Silpa-archa N,Kohli I,Chaowattanapanit S,et al.Postinflammatory (염기) 과색소 침착:포괄적인 개요:역학, 경로 발생, 임상 발표 및 비침습적 평가 기술 [J]다.J Am Acad Dermatol,2017,77 (4):591-605.
[10] 2013년 7월 1일에 확인함. Sies H,Stahl W.Nutritional protection 반대 skin 햇빛에 의한 손상 [J.Annu Rev Nutr,2004,24:173-200.
[11]Xuan Y,Yang Y,Xiang L,et al. 백반증의 발병에서 산화스트레스의 역할:A 범인 for melanocyte [J] 죽음이다.Oxid 의대Cell Longev,2022,2022:8498472.
[12]Decean H,Perde-Schrepler M,Tatomir C,et. modulation of the pro-inflammatory cytokines and matrix metalloproteinases produc러 졌 in co-cultivated 인간 각질세포와 멜라노사이트 [J. Arch Dermatol Res,2013,305 (8):705-714.
[13]Yuan XH,Jin ZH.melanogenesis의 Paracrine 조절 [J].Br J Dermatol,2018,178 (3):632-639.
[14]Bastonini E,Kovacs D,Picardo M.Skin pigmentation and pigmen-증인 장애:Focus on epidermal/dermal cross-talk[J].앤데르-마톨,2016,28 (3):279-289.
[15] 김M,Han JH,Kim JH 등 분비 frizzled-related protein 2 (sFRP2)는 멜라노제닉 자극제로 기능한다;sFRP2의 역할 uv로 유발된 과색소성 질환에서는 [J.J 투자Dermatol, 2016년, 136 (1):236-244.
[16]Jiang L,Huang J,Lu J,et. ganoderma lucidum polysaccharide IL-6/STAT3/FGF2 경로를 통해 kerlate-nocytes 및 섬유아세포의 paracrine 효과를 억제함으로써 melanogenesis를 감소시킨다 [J].J 셀 Physiol, 2019년, 234 (12):22799-22808다.
[17] Kovacs D,Cardinali G,Aspite N,et al.Role of fibroblast-derived (섬유세포유래) 태양 렌티고의 과색소침착을 조절하는 성장인자 (J). Br J Dermatol,2010,163 (5):1020-1027.
[18]. (영어) Serre C,Busuttil V,Botto JM.의 내적, 외적 규정 인간 피부 melanogenesis와 색소 침착 [J].Int J Cosmet Sci, (영문) 2018년, 40 (4):328-347.
[19]Fu C,Chen J,Lu J,et al. role of 염증인자 in melano-genesis (리뷰) [J.Mol Med 담당자, 2020년, 21 (3):1421년- 1430.
[20]Yoshida M,Takahashi Y,Inoue S.Histamine은 H2를 통한 protein kinase A 활성화에 의해 melanogene-sis와 형태적 변화를 유도한다 receptors in human normal 멜라 노 사이트다 [J다]J 투자 Dermatol, 2000년, 114 (2):334-342.
[21] 윤제, 김이, 권s 등 노화섬유아세포가 노화를 촉진한다 색소 침착:노인성 렌티고의 잠재적인 치료 표적. 테라노틱스,2018,8 (17):4620-4632.
[22] Kim Y, 강은 B, 김 JC, et al.Senescent fibroblast-derived GDF15는 피부 색소 침착을 유도합니다 [J.J Invest Dermatol, 2020년, 140 (12):2478-2486.
[23]Xu Z,Chen L,Jiang M,et al.CCN1/Cyr61은 사람의 표피 멜라노세포에서 Integrin alpha6beta1,p38 MAPK, ERK1/2 신호경로를 통해 melanogen-esis를 자극한다 [J].J Invest Der-matol,2018,138 (8):1825-1833.
[24]Lim HK,Suh DH,Lee SJ, 등의 히알루론산의 회춘 효과 injection on nasojugal 홈:장래 랜 덤 화 분할 얼굴 임상 제어 연구 [J].J Cosmet 레이저 Ther,2014,16 (1):32-36.
[25] HONG Wei-Song, QIAN Guo-pei, XU Ai-E.sodium vitrate 가 melanocytes의 생물학적 활성에 미치는 영향 (J.sodium vitrate 가 melanocytes의 생물학적 활성에 미치는 영향 (J.중국피부과학회지,2011,44(7):491-493.
[26]Muto J,Sayama K,Gallo RL,et al.Emerging evidence for the es-sential role of hyaluronan in cutaneous biology[J] (영어).J 피부과몰 Sci, 2019,94(1):190-195.
[27]Litwiniuk M,Krejner A,Speyrer MS,et al.Hyaluronic acid in-flammation and tissue regeneration[J].상처,2016,28 (3):78 -88.
[28] 타카베 P, 카르나 R, 라우할라 L, 외. 멜라노사이트 히알루로난 코트 단편화는 uvb로 유도된 TLR-4 수용체 신호를 강화시킨다 과 표현 of proinflammatory 중재자들 IL6, IL8 CXCL1, and CXCL10 via NF-kappaB 활성화다 [J다]J Dermatol 투자, 2019년, 139 (9):1993-2003.
[29] Marinho A,Nunes C,Reis S.Hyaluronic acid:의 주요 성분 치료 of 염증다 [J다] 생체 분자, 2021년, 1518년 (중종 11) 10월:중종반정.
[30] Campo GM,Avenoso A,Campo S,et al.의 DNA frag 감소-hyaluronic acid에 의한 침전과 hydroxyl radical 생성 및 chondroitin-4-sulphate in 철 게다가 ascorbate-induced oxidative 섬유아세포 배양에서 stress [J.자유 Radic Res,2004,38 (6): 601-611다.
[31] Cirillo N,Vicidomini A,Mccullough M,et al.A 히알루론산 기반 화합물은 산화성에 의해 유도된 섬유아세포 노화를 억제한다 in vitro에서 스트레스 및 in vivo에서 구강 점막염을 예방 [J].J 세포 physi-올,2015,230(7):1421-1429.
[32] Maytin EV다.히알루로난:단순한 주름필러 [J] 이상의 것.Glyco-생물학, 2016년, (6) 26일:553-559다.
[33] Bourguignon LY, Bikle D.Selective hyaluronan-CD44 signaling 피부 편평상피세포암 진행 중 miRNA-21발현을 촉진하고 비타민 D func-tion과 상호작용하여 자외선 조사 (J.Front Immunol,2015,6:224.
[34] Bourguignon LY,Wong G,Xia W,et al. 선택적 행렬 (히알루-로난) 과의 상호작용 CD44 and RhoGTPase signaling promotes keratinocyte functions and 극복 노화 epidermal [J] dys-기능이다.J Dermatol Sci,2013,72(1):32-44.
[35] Lagatta A, D'agostino A,Schiraldi C,et al.A biophysically-defined를 사용한다 hyaluronic 산 기반 화합물은 마이그레이션을 가속화합니다 stimulates the production of keratinocyte-derived neuromodulators다 [J다]Cell Adh Migr,2019,13 (1):23-32.
[36] Lee SG,Yoon MS,Kim DH,et. hyaluronan Oligosaccharides 개선 Rosacea-Like 개체 through 항 염증 그리고 표피 배리어 개선 효과 [J.Ann Dermatol,2020,32 (3):189-196.
[37] 후 L, 노무라 S, 사토 Y, 등 uvb로 유도된 calpro-tectin 매개 각질세포염증에 대한 차이-ential 분자량 히알루론산의 항염증 효과 [J/OL].J 피부과몰 Sci, 2022.[2022-06-10]다. https://www. jdsjournal다. com/article/S0923-1811 (22) 00158-X/fulltext.
[38] David-Raoudi M,Tranchepain F,Deschrevel B,et al.Differential effects of hyaluronan and its 섬유아세포의 조각:상처 치유와 관계.상처 수리 Regen,2008,16 (2):274 - 287.
[39] Ciccone V,Zazzetta M,Morbidelli L.Comparison of the effect of two hyaluronic acid preparations on fibroblast and endothelial cell functions related to angiogenesis[J] (영어).Cells,2019,8 (12):1479.
[40]Asparuhova MB,Kiryak D,Eliezer M,et al.Activity of two hyalu-ronan phrases on primary human oral fibroblasts (J).J Perio-dontal Res,2019,54(1):33-45.
[41] Quan T,Wang F,Shao Y,et al.Enhancing structural support of the의 약자 dermal microenvironment activates 생체 내 노화된 인간 피부의 섬유아세포, 내피세포 및 각질세포 [J.J Invest Dermatol,2013,133 (3):658-667.
[42] Radrezza S, 남작 G, Nukala 누군가, et al.Advanced quantitative proteomics to 평가 molecular effects of 사람의 진피 섬유아세포에 함유된 저분자 히알루론산 [J.J Pharm Biomed Anal,2020,185:113199.
[43] Loren CE,Dahl CP,Do L,et al. 저분자량 심근 hyaluronan in human 비대 심근 증다 [J다]세포, 2019년, 8 (2):97.
[44] Olsson M,Bremer L,Aulin C,et al.Fragmented hyaluronan은 없다 alarmin function 평가 in 관절염 관절 섬유 아세 포 그리고 chondrocyte 문화 [J]다. 타고 난 Immun, 2018, 24 (2):131~141 쪽.
[45]Zheng BW,Wang BY,Xiao WL,et al.Different molecular weight 히알루론산은 dnfb로 유도된 mouse 아토피 피부염과 lps로 유도된 RAW 264.7세포에서 염증 반응을 완화시킨다 [J].삶 Sci, 2022년, 301:120591다.
[46]Zhou J,Ling J,Wang Y,et al.Cross-talk between interferon-gam-ma and interleukin-18 in melanogenesis[J].J Photochem Photobiol입니다 B, 2016년, 163:133-143다.
[47] D'agostino A,La Gatta A,Stellavato A,et al.Potential of biofer-mentative unsulfated chondroitin and hyaluronic acid in dermal re-쌍 [J].Int J Mol Sci,2022,23 (3):1686.




