

 영어
영어 프랑스
프랑스 스페인
스페인 러시아
러시아 한국
한국 일본
일본[화학공학] 합성 리코펜 분말에 대한 자료입니다
Lycopene (C40H56) is a natural fat-soluble pigment of plant and microbial origin다. Chemically, it is a carotenoid composed of a straight-chain hydrocarbon with 11 conjugated double bonds and 2 non-conjugated double bonds [1]. Lycopene can effectively scavenge free radicals in the body and quench singlet oxygen. Its ability to quench singlet oxygen is 2 times that of β-carotene and 10 times that of α-tocopherol[2]. It can be used as an effective antioxidant to reduce the damaging effects of oxidative stress on cells. More and more studies have shown that lycopene has a protective or intervention effect on chronic diseases such as type 2 diabetes, malignant tumors and Alzheimer'의 질병이며, 따라서 식품, 화학공업, 의약 분야에서 많은 관심을 끈다.유엔 식량농업기구 (FAO), 식품첨가물위원회 (CCFA), 세계보건기구 (WHO) 에서는 리코펜을 a 급 영양소 [3]로 규정하고 있다.
천연 기능성 제품에 대한 인식이 높아지면서 기능성 식품 첨가물로서 리코펜에 대한 연구도 심화되고 있다.그러나 인체는 리코펜을 직접 합성할 수 없고 천연 채소와 과일 또는 장내 식물로부터만 얻을 수 있으며, 그 공급원과 양은 상당히 제한되어 있다.이 글에서는 리코펜 분말의 주요 구조, 생체 이용성 및 영향 요인, 미생물의 합성 경로, 리코펜을 생산하기 위한 세 가지 효모 균주의 합성 전략, 만성 질환 예방을 위한 리코펜 적용의 관계와 역할에 초점을 맞습니다.리코펜의 생산, 이용 및 기능성 탐구를 위한 이론적 기초를 제공한다.

리코펜과 그 유사체의 화학구조 및 생체이용가능성 1
리코펜 분자의 분자 구조에는 13개의 이중 결합이 있으며,이 중 11개는 공액결합으로, 리코펜은 불안정하고 빛, 산소, 산, 촉매 또는 기타 환경 변화의 영향으로 이성질화되기 쉽습니다.리코펜은 주로 올 e-이성질체 (올 트랜스 이성질체)와 z-이성질체 (시스-트랜스 이성질체)의 두 가지 형태로 존재한다 (그림 1). 과일과 채소에 함유된 천연 리코펜의 90% 이상은 열역학적으로 가장 안정적인 올 e-이성질체 형태로 존재한다 [4];그러나 연구에 따르면 인간의 혈청과 조직에 있는 리코펜의 50% 이상이 z-이성질체 [4]의 형태로 대사된다.일반적인 z-이성질체는 주로 5-cis, 9-cis, 13-cis, 15-cis 리코펜이다.연구에 따르면 5-cis 리코펜은 다른 유사체 [5]에 비해 항산화 능력이 높고 생체이용도 높은 것으로 나타났다.
따라서 5-cis 리코펜의 섭취는 all-E-lycopene보다 인체 건강에 더 유익할 수 있으며, 식품 및 제약 산업에 적용 가능성이 더 크다.최근 학자들은 고농도의 z-리코펜을 얻기 위한 열처리, 마이크로파 처리, 광조사, 전기분해 처리, 촉매 처리 등의 방법을 개발하기 위해 심혈을 기울이고 있다.그러나 이러한 방법에는 아직 개선의 여지가 있다.예를 들어, 가열 및 마이크로파 처리는 높은 온도로 인해 열화를 일으킬 수 있습니다;또한 광화학 처리는 올 e-이성질체의 전환으로 인해 열화를 일으킬 수 있다.비록 광감응제를 사용하면 리코펜의 광분해를 효과적으로 방지할수 있지만 광감응제 제거라는 난제를 가져다준다.마찬가지로 전해질이나 촉매와 같은 화학시약을 사용할 경우 독성물질을 제거하는 것 또한 큰 과제이다.
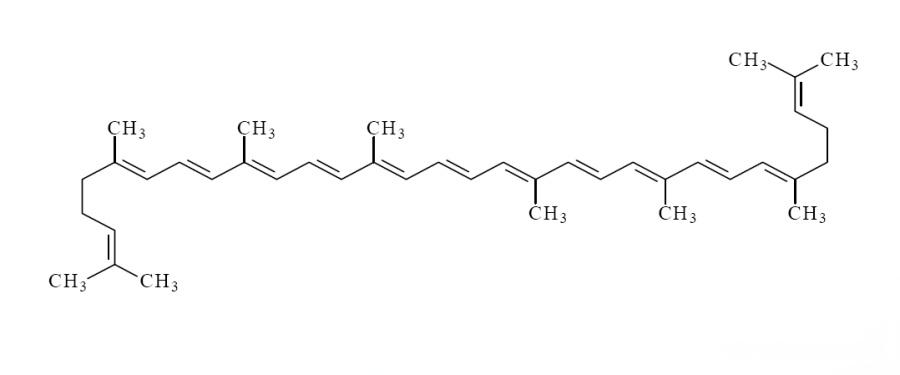
리코펜 분말의 생물학적 이용성에 영향을 미치는 두 가지 주요 이유가 있는데, 리코펜이 식품 매트릭스에서 완전히 방출되는지와 리코펜 의존성 지질 유화 및 미셀 형성 강도 [6] (그림 2). 리코펜의 극히 소수성 화학 구조로 인해 [7] 인체에 의한 과일과 채소에서 리코펜의 직접 흡수 및 이용률은 매우 낮다.그러나 식품 가공 중 열처리 등의 과정은 세포막을 손상시키고 조직 기질에서 리코펜의 분비를 촉진시켜 리코펜의 생체 이용성을 높일 수 있다.리코펜의 생체 이용성은 가공 방법에 따라 크게 달라지는데, 그 순서는 열처리 및 정제 유성 조제>가벼운 처리>생토마토 [8].리코펜의 효과적인 이용률을 더욱 높이기 위해 연구자들은 물리화학적 특성 및 체세포의 구조와 같은 특성을 바탕으로 전통적인 에멀젼, 나노에멀젼 운반체, 나노구조 지질 운반체 등의 리코펜 전달 시스템을 성공적으로 개발하였습니다 (그림 3). 이들 시스템은 활성 성분의 수용성 및 생체이용성을 높이기 위해"패키징"함으로써 리코펜의 생체이용성을 크게 향상시킬 수 있습니다,소화관의 악조건으로부터 보호하고, 흡수부위에 배출시켜 흡수가 더 잘 되도록 하는 것.
리코펜의 생합성 2
Natural lycopene is mainly derived from tomatoes and fruits such as grapefruit, melons, red guava, red carrots, and wolfberries. In addition, studies have confirmed that some microorganisms, including bacteria, fungi and algae, can accumulate lycopene under specific physiological conditions [9]. For example, the inactivation of lycopene cyclase leads to the interruption of the carotenoid pathway, which helps lycopene accumulate in Blakeslea trispora [10]. The Codex Alimentarius Commission (CAC) has approved three sources of lycopene: tomato extract, chemical synthesis, and Blakeslea trispora extract.
Among them, the tomato extraction method mainly uses vegetable and fruit raw materials rich in lycopene, which are extracted efficiently using various extractants. The advantage of this method is that it can achieve high-quality natural lycopene production in batches, but this method is susceptible to external factors such as the species, origin, and harvest season of the raw materials, which can affect the yield. In addition, large amounts of waste residue, waste liquid, and waste gas are generated during industrial production, which results in high comprehensive treatment costs. The chemical synthesis method is relatively mature, with mild reaction conditions, high recovery rates and low costs. It is currently the main technology for the industrial production of lycopene. However, lycopene has many C=C double bonds, making it difficult to control stereoselectivity. The reaction process is complex and has high technical requirements. There is also the safety issue of organic solvent pollution from the chemical reagents left over from the reaction. In recent years, with the analysis by scientists of the biosynthetic pathway of natural 리코 펜 가루 and the great progress in modern microbial genetic engineering, other microorganisms (such as Saccharomyces cerevisiae, Pichia pastoris and Yarrowia lipolytica) can also be used as hosts for lycopene production. Because they have the incomparable advantages of no seasonal restrictions, high yield and a single product, they provide a new way of thinking for the large-scale industrial production of lycopene and have attracted the attention of researchers and the food and pharmaceutical industries.

2.1리코펜의 생합성 경로
생물에서 리코펜은 주로 두 가지 생합성 경로를 통해 합성된다:메발로네이트 (mevalonate, MVA) 경로와 2-methyl-D-erythritol-4-phosphate (2-methyl-D-erythritol-4-phosphate, MEP) 경로.그 중 진핵생물은 주로 라이코펜과 그 유도체를 MVA 경로를 통해 합성하고, 원핵생물은 MEP 경로를 통해 합성하는 경우가 많다.두 가지 생합성 경로 모두 body&에서 생성되는 glyceraldehyde-3-인산염 (G3P)을 사용합니다#39;의 당 대사, isopentenyl pyrophosphate (isopentenyl pyrophosphate, IPP) 및 이의 이성질체 3,3-dimethylallyl diphosphate (DMAPP) 및 기타 중간 분자와 같은 중간 분자를 합성하기 위해 일련의 2차 대사 효소를 촉매하기 위해.이어서 IPP와 DMAPP은 효소에 의해 응집되고 변형되고 길어져 최종적으로 리코펜을 합성한다 (그림 4).
리코펜 합성을 위한 효모의 대사공학 2.2
자연에서는 Rhodotorula glutinis, Rhodotorula graminis 및 Phaffia rhodozyma와 같은 효모들은 자율적으로 카로티노이드 천연 생성물을 합성할 수 있지만, 합성된 생성물의 양과 생물학적 활성은 종종 산업 생산의 요구를 충족시키지 못한다 [11].그러나 일반적으로 사용되는 사카로마이세스 세레비지애 (Saccharomyces cerevisiae), 피치아 파스토리스 (Pichia pastoris), 야로위아 리폴리티카 (Yarrowia lipolytica)와 같은 공업용 발효 효모들은 안전성이 높고, 성숙한 유전자 변형 도구를 가지고 있으며, [12] 리코펜의 연구 및 생산을 위해 유전자 변형이 되었다.S. cerevisiae와 Y. lipolytica에서 완전한 대사체계를 형성하지 못하고, 카로티노이드 합성의 대사과정이 geranylgeranyl diphosphate (GGPP) 단계에서 멈추는 점 [13]에 대해 학자들은 리코펜을 생산하는 효모 균주를 구축하기 위한 다양한 전략을 제시하고 있으나 (표 1), 최종 리코펜 역가와 수율을 향상시키는 것은 여전히 큰 과제로 남아 있다.P. pastoris와 Y. lipolytica에 비해 S. cerevisiae에 대한 전략이 더 많이 보고되고 있으나, S. cerevisiae에 대한 체계적인 공학적 연구를 통해 높은 리코펜 생산을 얻을 수 있는 연구는 적은 실정이다.heterologous pathway 성분의 원천과 pathway의 효율은 S. cerevisiae에서 lycopene 생산에 핵심이며 [14], 낮은 수율은 내인성 경로와 이인성 경로 간의 조화가 잘 이루어지지 않아 발생하는 것이 가장 크다.
따라서, S. cerevisiae 자체의 적응성과 이형 경로를보다 자세히 탐색하기 위해, SHI B 등 [6]은 리코펜의 생합성에 관여하는 박테리아, 효모, 곰팡이, 조류, 식물 등의 다양한 원으로부터 crtE (encoding GGPP synthase), crtB (encoding octahydro-lycopene dehydrogenase) 및 crtI (encoding octahydro-lycopene dehydrogenase;스크리닝 된 유전자의 조합은 내인성 및 외인성 대사 경로 간의 불균형으로 인한 주요 단계의 손실을 방지한다;내인성 바이패스 유전자를 제거하면 전구체 아세틸 코엔자임 A (acetyl coenzyme A, acetyl-CoA) 공급과 균형 잡힌 NADPH 이용이 증가하며, 순수한 포도당 유도 시스템이 달성되었고, 가장 높은 수율을 갖는 균주인 BS106 (리코펜 수율 3.28 g/L)을 제조하였다.이 균주는 S. cerevisiae&의 호환성 향상을 위한 참고를 제공한다#39; 내인성 배경으로 가치 있는 물질을 생산하는 s 이형 경로.현재, 리코펜을 포함한 이소 프레 노이드 화합물의 미생물 생산은 두 가지 잠재적인 도전에 직면해 있다:천연 MVA 또는 MEP 경로는 보조 인자에 의해 제한된다;그리고 대부분의 긴사슬 이소프레노이드 화합물은 소수성 때문에 주로 한정된 공간에 저장되는데, 이렇게 되면 대규모 축적을 못하게 된다 [15].
두 가지 문제를 해결하기 위해 LUO Z S 등 [16]은 isopentenol을 IPP로 직접 전환하고 MVA 경로를 강화하며 IPP 및 다운스트림 제품의 플럭스를 증가시키는 IUP (isopentenol utilization pathway)을 도입했다 [14].IUP과 높은 소수성을 결합하면 Y. lipolytica 가 지질 용해성 이소펜테닐 화합물의 축적에 더 도움이 되는 지질 생성 생물로 전환된다.이러한 전략은 상업적 목적으로 널리 사용될 수 있습니다.P. pastoris는 중요한 상업적 이점 또한 가지고 있기 때문에 카로티노이드 물질 생산지로 선정되었다.P. pastoris는 세포질이 높고 에탄올을 축적하지 않고 다양한 종류의 유기물을 탄소원으로 사용하지 않고도 S. cerevisiae와 같은 다른 효모보다 높은 밀도로 자생이 가능하다.따라서 BHATAYA A 등 [17]은 P. pastoris에 처음으로 대사공학 기술을 적용하여 두 개의 플라스미드를 설계하고 제작하였다:pGAPZB-EpBPI*P 플라스미드는 표적이 되는 superoxide dismutase를 암호화하고, pGAPZB-EBI* 플라스미드는 표적이 되지 않는 효소를 암호화한다.이 두 플라스미드가 P. pastoris로 변형된 후, pGAPZB-EpBPI*P 플라스미드를 포함하는 P. pastoris v 형 클론의 고생산성 라이코펜 생산 균주를 스크리닝할 수 있었으며, P. pastoris를 이용한 라이코펜 생산 개발의 기초를 마련할 수 있었다.
합성생물학, 단백질공학, 대사공학의 급속한 발달로 유전자조작 효모는 리코펜의 생산효율을 향상시켰을 뿐만 아니라 저렴한 기판의 활용도를 높여 생산비용을 더욱 절감시켰다.합성 미생물은 의심의 여지없이 천연 제품의 이형 합성에 새로운 옵션을 제공 할 것입니다.
리코펜의 항산화 생리활성 3
악성 종양과 산화 스트레스 등 만성질환의 발병과 진행 사이에 연관성이 있다는 연구 결과가 나왔다.리코펜은 천연 항산화제로서 산화적 스트레스로 인한 해를 감소시키는 효과가 있다.리코펜 분말의 주요 항산화 활성은 과산화수소, 이산화질소 및 히드록시기 등의 활성산소에 작용하여 단백질, 지질 및 DNA의 산화를 억제하는 것입니다.리코펜이 산화제나 활성산소에 노출되면 이중결합이 쪼개지거나 증가해 폴리엔 사슬이 파괴될 수 있다.활성 물질과 리코펜의 가능한 반응은 [32] 다음과 같다:첨두의 형성, 활성산소로의 전자 이동 및 대립형질에서 수소의 추출 (그림 5). 다음 설명은 여러 만성 질환과 산화 스트레스의 관계와 리코펜이 만성 질환을 유발하는 돌연변이를 어떻게 억제하는지에 초점을 맞추고 있다.
종양세포는 보통 반응성 산소종 (ROS) [33]이 지나치게 높고 산화 스트레스를 경험한다.ROS는 신호전달에 핵심적인 역할을 하는 세포의 정상적인 대사산물이다.종양 세포에서 높은 수준의 ROS는 종양 세포 성장, 증식, 침입, 혈관 생성 및 전이 [34]와 같은 다양한 종양 생성 단계에 관여한다.리코펜과 시스플라틴이 인간 자궁경부암 (HeLa) 세포의 성장을 억제하는 시너지 효과가 있다는 것이 밝혀졌다.리코 펜으로 치료 됐 는지 세포의 72 시간 생존 률 (10 μ mol/L)과 라틴 (1 μ mol/L) 혼자 65.6%었고 71.1%, 셀을 각각, 그리고 생존능력 조합 후 37.4%에 감소하였다.또한 대조군과 비교하여 라이코펜이 처리된 세포군은 nuclear factor E2-related factor (NRF2)의 발현이 증가하였으며, 시스플라틴을 처리한 세포군만 투여한 경우보다 병합투여군의 NRF2 수치가 유의하게 높았다.이러한 결과는 리코펜이 산화 스트레스를 매개하기 위해 NRF2를 활성화함으로써 항암 효과를 발휘할 가능성이 있음을 나타낸다 [35] (그림 6).
비정상적인 NRF2 신호 조절은 많은 산화 스트레스 관련 질병과 관련이 있다.NRF2의 활성화는 주로 NRF2 신호를 매개로한 항산화 효소의 유도를 통해 항산화 능력을 유도하고 병리를 완화할 수 있는 방법으로 여겨진다.또 다른 연구에서, 그것은 리코 펜 핵 요인의 활성화를 억제 할 수 있 다는 것을 발견 κ-light-chain-enhancer의 활성화 B 세포 (NF-κ B) 및 NF의 표현-κ B 대상 유전자 (cIAP1, cIAP2 및 survivin)을 줄 임 으로써 세포 내 및 미 토 콘 드리아 ROS 수준, 유도하는 췌장암에서 PANC-1 세포 사멸 했다.이러한 연구 결과는 리코펜 보충제가 잠재적으로 췌장암을 예방할 수 있음을 시사한다 [36].
염증은 몸 's 자신의 방위 대응.정상적인 신체 균형에서 염증은 세포 손상을 일으키는 초기 요인을 제거하고, 손상과 염증으로 인해 괴사된 세포와 손상된 조직을 처분하고 조직 복구를 수행하는 역할을 한다.이 자연적인 반응인 급성 염증은 모든 고등 척추동물들이 사용하는 핵심적인 생존 메커니즘이다 [37-38].그러나 급성염증이 해결되지 못하면 만성염증으로 이어질수 있으며 파괴적인 과정으로 될수 있다.손상된 조직은 친염증성 사이토카인과 다른 생물학적 염증 매개체를 체내에 & 방출한다#39;s 순환계, 따라서 저급조직염증을 전신염증 [39]으로 변형시킨다.또한 자가면역질환이나 자극물질에 장기간 노출될 경우 전신적인 염증상태에 이르기도 한다.과도한 염증반응은 신체&에 악영향을 미치게 됩니다#39;s 가 회복되고, 염증 침윤 [40]의 장기간의 자극으로 세포가 암이 될 수 있다.연구들은 리코 펜 미 토 콘 드리아 장애를 개선 할 수 있다고 보고 lipopolysaccharide에 의해 유도 된 쥐의 뇌와 간, 겁 TNF-a 질리의 표현 수준을 줄이고, IL-1 β 그리고 IL-6, 그리고 neuroinflammation을 완화하고 간염 [41]이다.
4 결론 및 전망
This paper provides a systematic review of recent research progress on the structure, bioavailability, heterologous microbial synthesis strategies, and protection against oxidative stress in chronic diseases of lycopene powder다.리코펜은 카로티노이드 계열에 속하며, 항산화 능력은 건강에 상당한 이점이 있습니다.이 특성은 식품 제형에서의 사용에 큰 관심을 불러 일으켰다.이 화합물을 사용하기 위해서는 추출 및 유지 공정이 리코펜의 안정성 및 생체 이용성에 영향을 미치는 인자를 충분히 고려하도록 해야 매우 효과적이고 활용성이 높은 기능성 제품을 얻을 수 있습니다.
생리활성물질에 대한 전통적인 추출기술로는 기계적 추출법과 초음파 추출법, 안전한 유기용매를 이용한 추출법 등이 많이 이용되고 있다.그러나 다양한 분야의 발전으로 인해 리코펜 추출의 가능성이 큰 고전단 혼합, 고압 균질화, 미세유체 처리 등 새로운 대체 방법이 등장하고 있다.또한, 초미세 분쇄는 리코펜의 추출률을 향상시킬 뿐만 아니라 식품 등급 용매에도 좋은 선택인 새로운 옵션입니다.리코펜 보호 측면에서 리코펜 전달 시스템은 리코펜의 체내 활용도를 보호하고 향상시킬 수 있는 대체 방법이 되었습니다.나노 에멀젼 운반체, 나노 구조 지질 운반체, 하이드로겔, 리포좀의 개발은 리코펜의 보호를 향상시키는 좋은 선택입니다.
In addition, the use of industrial yeast as a host cell to produce lycopene is also a brand new idea. Yeast that does not have a lycopene powder synthesis pathway can become a lycopene-producing strain by introducing genes from an external source. This strategy improves the production efficiency of lycopene and reduces production costs. Under the premise of achieving high lycopene yields, scholars can also develop other effective methods to synthesize other high-value carotenoids.
참조:
[1]SOUKOULIS C, BOHN T. 「 A comprehensive overview on the micro-and nano-technological encapsulation advances for enhancing the chemical stability and bioavailability of carotenoids 」 (영어).식품영양학의 비판적 고찰 (Critical Reviews in Food Science and Nutrition),2018,58(1):1-36.
[2]PRZYBYLSKA S. Lycopene-a bioactive carotenoid offers multiple health benefits:a review[J].국제학술지 한국식품과학&기술, 2020년, 55 (1):11-32다.
[3] 량 X P, 마 C C, 얀 X J 외.리코펜의 생체활성, 대사, 안정성 및 전달체계에 대한 연구의 발전.식품과학&의 동향기술, 2019년 93:185-196다.
[4] 클린턴 S K, 에멘하이저 C, 슈워츠 S J 외.시스-트랜스 리코펜 이성질체, 카로티노이드, 그리고 레티놀린은 인간의 전립선 [J.암 역학 바이오 마커&방지, 1996년, 5 (10):823-833.
[5] 혼다 M, 카게야마 H, 히비노 T 등.자연적으로 발생하는 z-이성질체 가속 촉매를 이용하는 Paracoccus carotinifaciens 로부터 효율적이고 친환경적인 카로티노이드 추출법 [J].프로세스 생화학, 2020년, 89:146-154다.
[6] 시아파 B, MA T, YE Z L, 외. lycopene 과잉생산을 위한 Saccharomyces cerevisiae의 체계적 대사공학 (J.한국식품체-미스트리 2019년,67(40):11148-11157.
[7]SHARIFFA Y N, TAN T B, ABAS F 등.리코펜 나노분산 생성:유화제의 효과 (Producing a lycopene nanodispersion:the effects of emulsifiers[J])식품바이오제품가공,2016,98:210-216.
[8]HONEST K N, ZHANG H W, ZHANG L. Lycopene:생물이용성 및 생물활성 특성에 대한 이성질화 효과 [J.Food Reviews International,2011,27(3):248-258.
[9] 펑 L R, 치앙 W, 유 X B 외.Blakesleatrispora의 lycopene 생성과 지방산 조성에 미치는 외인성 지질 및 냉적응의 영향 (J.다 표현, 2019년, 9 (1)이다.
[10]MEHTA B J, CERDAOLMEDO E. Mutants of carotene production in Blakesleatrispora[J].「 Applied Microbiology and Biotechnology 」,1995,42(6):836-838.
[11]리 C J, 장 N, 송 J 외.적색 효모 Sporidiobolus pararoseus의 단일 desaturase 유전자는 phytoene의 4단계 및 5단계 탈수소화를 모두 담당한다 [J].유전자, 2016년,하이 (1):169-176다.
[12] 손링, 왕준화, 장위 등.고효율 합성 리코펜 효모 균주 제작 [J.중국생명공학저널 2020, 36(7):1334-1345.
[13]VERWAAL R, WANG J, MEIJNEN J P 등.고위급 Saccharomyces cerevisiae에서 carotenogenic gene 과의 연속적인 형질전환에 의한 beta-carotene 생산 Xanthophyllomyces dendrorhous[J.적용 되고 2007년 환경 미생물학, 73 (13):4342-4350다.
[14]MA T, SHI B, YE Z L 등.리코펜의 고수율 생산을 위한 Saccharomyces cerevisiae의 체계적 대사공학을 이용한 지질공학 (Lipid engineering)과 Saccharomyces cerevisiae (J.대사 공학, 2019년, 52:134-142.
[15] 징 Y W, 구 F, 장 S J 등.최근 산업용 효모를 이용한 리코펜의 생물학적 합성에 관한 연구 (Recent advances on biological synthesis of lycopene by using industrial yeast[J.산업 &공학화학연구,2021, 60(9):3485-3494.
[16] 루오 Z S, 류 N, 라자르 Z 등.에 의해 Yarrowialipolytica에서 isoprenoid 합성 향상 isopentenol 이용 경로를 표현하고 세포 내 소수성을 조절함 [J.대사 공학, 2020년, 61:344-351.
[17]BHATAYAA, SCHMIDT-DANNERT C, LEE P C. meta-bolic engineering of Pichia pastoris X-33 for lycopene production[J].2009년 과정 생화학, 44 (10):1095-1102.
[18]리 X, 왕즈 X, 장 G L 등.경로 및 차체대사 최적화를 통한 Saccharomyces cerevisiae의 리코펜 생산 개선 [J.화학공학연구 2019년, Chemical Engineering Science, 193:364-369.
[19]HONG J, PARK S H, KIM S, 외.효소공학에 의한 Saccharomyces cerevisiae의 lycopene의 효율적 생산과 막의 유연성 및 NAPDH 생산 증대 [J.한국미생물학회지 Applied Microbiology and bio-technology,2019,103(1):211-223.
[20] 알셰흐리와, 가달란 오, 에드리스 S 외.pESC-LEU와 pTEF1/Zeo 벡터를 이용한 Saccharomyces cerevisiae에서 lvcopene의 과잉생산을 위한 mep pathway의 메타-볼릭 공학 (Meta-bolic engineering of mep pathway for overproduction of Saccharomyces cerevisiae)응용생태학 및 환경 세서치,2020,18(4):5279-5292.
[21] 장 Y, 치우트 Y, 장 J T 등.라이코펜의 생산 증진을 위한 합성 효모의 체계공학 (Systematical engineering of synthetic yeast for enhanced production of lycopene[J])2021년 생명공학,, 8, (1):14이다.
[22] 수 X, 류 J, 루 Y L 등.for Saccharomyces cerevisiae의 경로 공학 효율적인 리코 펜 생산 [J]이다.바이오공정 및 바이오시스템공학, 2021,44(6):1033-1047.
[23] 비안 Q, 저우 PP, 야오 Z 외.temporospatial pathway control에 의해 가능하게 된 S. cerevisiae의 lutein의 Heterologous biosynthesis (J.대사 공학, 2021년, 67:19-28.
[24] 수 B L, 양 F,리 A Z 외.업스트림 활성화 서열은 Saccharomyces cerevisiae에서 위치 효과에 대한 이형 경로의 염색체 조절을 위한 절연체로 기능할 수 있다 [J].응용생화학과 생명공학 (Applied Biochemistry and Biotechnology),2022,194(4):1841-1849.
[25] 수 B L, 라이 P X, 양 F, 외.부제:a balanced acetyl coenzyme a metabolism in Saccharomyces cerevisiae for lycopene production through rational and evolutionary Engineering [J]한국농식품화학학회지 2022,70(13):4019-4029.
[26] 아라야-가레이 J M, 페이주-시오타 L, 로사-도스-산토스 F 등.lycopene 및 beta-carotene 생산을 위한 새로운 PichiapastorisX-33 균주의 구축 [J.한국미생물학회, Applied Microbiology and Biotechnology,2012,93(6):2483-2492.
[27] 장 X, 왕 D, 듀안 Y 등.대사적으로 조작된 Pichiapastoris에 의한 리코펜의 생산 [J.생명공학,Biotechnology, and Biochemistry,2020,84(3):463-470.
[28] 류 D, 류 H, 키 H 등.지방성 테르펜 합성을 증진시키기 위해 효모 키메릭 통로를 구성함.ACS Synthetic Biology,2019,8(4):724-733.
[29] 장 XK, NIE MY, 첸 J 외.crt 유전자의 다코피 통합체와 AMP deaminase의 공동 발현은 Yarrowia lipolytica에서 리코펜 생성을 향상시킨다.한국생명공학회지 2019년, 제289회:46-54.
[30] 제아잔틴의 생합성을 위한 카로테나이드를 생성하지 않는 효모 Yarrowialipolytica의 대사공학 (XIE Y X, CHEN S L, XIONG X C.프론트 마이크로바이올,2021, 12:699235.
[31] 류 X Q, CUI Z Y, SU T Y 등.게놈의 식별을 개발을 위한 통합 사이트 CRISPR-based 유전자 발현 툴킷에서 Yarrowialipolytica다 [J다]Microbial Bio-technology,2022,15(8):2223-2234.
[32]MAGNE T M, BARROS A O D S D, FECHINE P BA 외.리코펜은 암과 염증 치료를 위한 다기능 플랫폼으로 [J.Revista Brasileira De Farmacognosia-Brazilian Journal of Pharmacognosy,2022, 32(3):321-330.
[33]LIBBY P, BURING J E, BADIMON L 외.Athero-경화증 [J]다.Nat RevDis Primers,2019,5(1):56.
[34] 갈라다리 S, 라흐만 A, 팔리찬디 S 외.Reactive oxygen species and cancer paradox:촉진 또는 억제?[J].한국활성산소학회지 2017년, Free Radical Biology and Medicine, 104:144-164.
[35] 악테페 오흐, 사힌 T K, 구너 G 외.리코펜은 표적 nuclear factor-kappa B (NF-kappaB) 경로를 통해 자궁경부암 세포를 시스플라틴에 감응시킨다 [J].터키의학저널,2021,51(1):368-374.
[36]JEONG Y, LIM J W, KIM H. Lycopene은 reactive oxygen species-mediated NF-kappaB 신호전달을 억제하고 췌장암세포에서 apoptosis를 유도한다 [J].영양분, 2019년, 11 (4):762.
[37]TODORIC J, ANTONUCCI L, KARIN M. Targeting 염증 in cancer prevention and therapy[J].암예방연구,2016,9(12):895-905.
[38] 메드지토프 (MEDZHITOV R.)-인플레암의 기원과 생리적 역할 (J.자연, 2008년 454 (7203):428-435다.
[39] 아울 셀반 P, 파드 M T, 탄 W S 등.염증에서 항산화제와 천연 제품의 역할.「 산화의학과 세포장수 」,2016,(13):5276130.
[40] 싱엔, 베이비 D, 라즈구루 JP 외.염증과 암 [J.Annals of African Medicine,2019,18(3):121-126.
[41]WANG J, ZOU Q H, SUO Y 등.리코펜은 간뇌축에서 인슐린 저항성과 미토콘드리아 기능 장애를 개선함으로써 전신적인 염증유발성 시냅스 기능 장애를 개선한다.음식 &함수, 2019년 10 (4):2125-2137다.




