

 영어
영어 프랑스
프랑스 스페인
스페인 러시아
러시아 한국
한국 일본
일본리코펜 분말의 생산방법에 관한 연구
리코 펜 is an important carotenoid that belongs 을이terpene family 의tetraterpenoids. It is found 에서nature mainly 에서tomatoes, tomato products, 그리고fruits such as watermelons 그리고grapefruits. It is 이main pigment in ripe tomatoes. Lycopene is a strong antioxidant with physiological functions such as anti-oxidation, anti-cancer 그리고lowering blood lipids. It is widely us에드in health foods, medicine, cosmetics 그리고other fields [1-2]. At present, 리코 펜has been widely used as a nutritional supplement 그리고coloring agent in many countries [3]. The global cumulative sales 의리코 펜raw materials are increasing year 에 의해year, 그리고the market prospects are broad.
리코 펜 가루생산mainly relies on two methods: plant extrac졌그리고화학synthesis. The plant extraction 방법is limited 에 의해the season, 그리고the long plant growth cycle and low product content cannot ensure intensive and large-scale 생산의the product. The chemical synthesis method has problems such as chemical reagent residues, multiple isomeric forms, and environmental pollution. The biotechnology synthesis method has the advantages 의low cost, short cycle, stable supply, and environmentally friendly and sustainable development. In recent years, it has attracted more and more attention from researchers.
현재, 라이코펜 분말의 생명공학적 합성에 관한 연구는 미생물 숙주세포의 선택의 다양화, 경로를 변환하기 위한 대사공학의 연구 및 혁신, 생명공학에서 합성한 라이코펜의 수율을 현저히 향상시킨 발효공정 및 증폭기술의 탐구 등 상당한 진전을 이루었다.그러나 아직까지 대부분의 연구는 단일 기술 분야의 획기적인 발견에 집중되어 있으며, 라이코펜의 생명공학적 합성에 대한 체계적인 연구와 요약은 상대적으로 미흡한 실정이다.
따라서 본 논문에서는 라이코펜의 물리, 화학적 특성과 현재 생산기술을 살펴보고, 합성생물학을 대표하는 생명공학을 이용한 라이코펜의 생합성에 관한 연구결과를 계통별 발효방법 비교와 정확한 라이코펜의 정량방법을 중심으로 체계적으로 요약하고, 생명공학을 이용한 라이코펜 생산의 문제점과 향후 연구방향을 제안하였다,생명공학을 이용한 리코펜의 산업적 생산과 생명공학을 이용한 기타 고부가가치 천연물의 생합성에 참고자료를 제공하는 것을 목적으로 한다.
1. 리코펜 분말의 물리적, 화학적 특성 및 응용
리코펜은 사염화화합물, 불포화 알케닐 화합물, 산소 원자를 포함하지 않는 카로티노이드이다.리코펜은 분자식이 C40H56 이고 상대 분자량은 536.85이다.분자구조에서 11개의 공액이중결합과 2개의 비공액이중결합을 가지며, cis-트랜스 이성질체 에도 종종 존재한다.자연계에서 천연 리코펜은 주로 올 트랜스 (all-trans) 이며, 시스는 매우 적은 양을 가지고 있다.
Lycopene powder is a fat-soluble pigment that is insoluble in water, but soluble in lipids and non-polar organic solvents. Its molecular structure contains a chromophore, which corresponds to a unique absorption region in the ultraviolet-visible absorption spectrum. The color depth varies from orange-yellow to dark red depending on the concentration 의lycopene, and may slightly change with the solvent. For example, 리코 펜crystals dissolved in sunflower oil appear a visible dark red, while dissolved in petroleum ether appear yellow. Due to the relatively large number 의double bonds in the molecule, lycopene is very reactive and prone to oxidation and structural isomerization reactions under light, oxygen, and high temperature conditions, which can lead to a decrease in physiological activity [4]. Therefore, when lycopene is extracted, antioxidants such as vitamin C, vitamin E, 2,6-di-tert-butyl-4-methylphenol (BHT), and tert-butylhydroquinone (TBHQ) are often added [5].

과는 달리β-carotene, lycopene does not have the pro-active properties 의vitamin A, so its 응용 프로그램was not valued in the early days. However, in recent years, as the physiological functions 의lycopene have gradually become better known, its application has become more widespread. Lycopene is a powerful antioxidant that can scavenge oxygen free radicals in the human body and quench singlet oxygen. Its antioxidant capacity is about 100 times that 의vitamin E and twice that of 베 타 카로 틴[6-9]. It has also been shown to have anti-tumor, prevent prostate disease and reduce the risk of cardiovascular disease [10-11]. It is widely used in cosmetics, health products and food. Lycopene has currently obtained the “novel food” approval of the European Union and the “generally recognized as safe” (GRAS) status in the United States. With the improvement of people'의 생활수준과 건강에 대한 강조가 갈수록 높아짐에 따라 미국은 라이코펜 판매량이 매년 35%의 속도로 성장할 것으로 예측한다.따라서 리코펜의 효율적인 생합성 기술은 시장 응용 가치가 크다.
리코펜 파우더의 생산 방법 2
2.1리코펜 분말 생산 방법 비교
현재는 3개가 있다methods 을생산lycopene: plant extraction, chemical synthesis, and biosynthesis. The plant extraction method mainly involves extracting and purifying lycopene from ripe plant fruits such as tomatoes. However, this method is affected 에 의해various factors such as region, season, tomato variety, and maturity, and is therefore unstable. In China, lycopene is mainly extracted from tomatoes grown in Xinjiang (with long days of sunshine). However, the lycopene content in tomatoes is very low, generally only 20 mg/kg, and even in the tomato skin, where the content is higher, it is less than 0.4 g/kg [12].
추출 비용이 높고, 추출물에 다른 카로티노이드가 들어 있는 경우가 많아 제품의 순도에 영향을 미친다.그리고 그 함량이 적기 때문에 추출과정에서 유기용매를 다량 소모하여 환경오염에 더 큰 영향을 미친다.화학적 합성방법은 주로 옥타트리에네다이얼과 트리페닐포스핀 클로라이드 또는 트리페닐포스핀 황화물의 Wittig 반응을 통해 리코펜을 합성한다 [13].화학 합성 방법은 높은 수율 (65% 이상), 저렴하고 재활용 가능한 원료, 온화한 반응 조건의 특성을 가지고 있습니다.화학합성법은 수율이 높고 원가가 저렴하지만, 리코펜 구조에 이중결합이 많아 이성질화하기 쉽고, 제품에 용매잔류물이 포함될 수 있어 안전성에 대한 위험이 있다.생합성법은 설탕, 물엿, 무기염 등 풍부하고 쉽게 구할 수 있는 원료를 이용해 미생물이 라이코펜을 발효 · 생산하는 과정을 말한다.미생물발효법은 식물추출법의 안전성 (둘다 생물물질대사에서 자연적으로 추출한 것으로 인공적으로 합성하지 않음)이 있을 뿐만 아니라 화학합성법의 원가가 저렴하고 대량생산이 가능한 장점이 있다.향후 라이코펜 생산을 위한 이상적인 방법으로 평가되고 있다.

2.2리코펜 분말의 생합성 경로
리코펜은 다른 테르페노이드와 유사한 4 테라페노이드 화합물이다.이것의 생합성을 위한 일반적인 전구체는 두 개의 이소펜테닐 단위 IPP (이소펜테닐 피로인산염)와 DMAPP (디메틸알릴피로인산염) 이며, 이들은 서로 이성질체이다 [14].현재 자연계에서 IPP와 DMAPP을 합성하는 방법은 두 가지가 있는데, 하나는 원핵생물과 식물에서 MEP (2-methyl-erythritol-4-phosphate) 경로이고, 다른 하나는 진핵생물에서 MV한(mevalonate) 경로이다.
원핵생물과 식물에서는 MEP (2-methyl-erythritol-4-phosphate) 경로를, 진핵생물에서는 MVA (mevalonate) 경로를 이용한다.
MEP 경로는 피루브산과 3-포스글리세레이트를 출발기질로 사용하여 IPP와 DMAPP을 합성하고 [15], MVA 경로는 아세틸 코엔자임 A를 출발기질로 사용하여 7단계의 효소반응 [16]을 통해 IPP와 DMAPP을 합성한다.MEP 경로에 비해 MVA 경로는 더 일찍 연구되었고 반응 메커니즘도 더 철저하다.리코펜의 생합성 경로는 두 가지 모듈로 나눌 수 있습니다.업스트림 모듈은 전구물질인 IPP와 DMAPP을 합성하는 과정이고, 다운스트림 모듈은 IPP와 DMAPP 으로부터 라이코펜을 합성하는 과정이다 (개요는 그림 1 참조).IPP와 DMAPP은 isopentenyl transferase의 작용으로 단계적 응결반응을 거쳐 GGPP를 생성하고, GGPP (geranylgeranyl pyrophosphate)는 octahydro-lycopene synthase (phytoene synthase, CrtB)의 작용으로 octahydro-lycopene dehydrogenase (phytoene desaturase, CrtI)의 작용으로 octahydro-lycopene으로 전환된다.
리코펜을 합성하는 미생물 2.3
현재 라이코펜을 생성하기 위해 발효하는 미생물로 알려진 것으로는 라이코펜 자체를 합성할 수 있는 판토에아 (pantoea), 트리스포라 (Blakeslea trispora), 대사작용을 하는 효모 (yeast), 야로위아 폴리티카 (Yarrowia lipolytica), 대장균 (Escherichiacoli) 등이 있다.그들 중, Blakeslea trispora 연구 되어 왔 더 [17] 그리고은 또한의 유일한 산업 생산 달성 할 수 있는 변종 β-carotene다.리코 펜, 중급 제품의 합성에 β-carotene, 중에 리코 펜 cyclase 억제 제를 추가 하여 축적 된 할 수 있 발효 과정이다.여러 연구에 따르면 Blakeslea trispora에 의한 리코펜의 생산은 지속적으로 개선되었으며, 가장 많이 보고된 리코펜 생산량은 3.4 g/L이다 [18].그러나 트리코테센 곰팡이는 하위배양 과정에서 변성이 일어나기 쉬워 산출량이 불안정하다.게다가 성장 주기가 길면 생산성이 떨어지고, 생산 중에 억제제를 넣어야 하기 때문에 트리코테센 곰팡이 [19]로 리코펜을 발효시키는 과정도 크게 제한된다.
리코펜의 생명공학적 합성에 관한 연구 3
3.1리코펜을 합성하기 위한 주요 미생물의 공학적 개질
대장균은 테르페노이드의 이형 합성을 위해 가장 일반적으로 사용되는 미생물 숙주 중 하나이다.명확한 유전적 배경, 빠른 세포 성장, 풍부한 유전자 조작 도구 등의 장점은 대장균을 산업 제품 개발에 이상적인 숙주 플랫폼으로 만들어줍니다.일부 학자들은 대사공학과 합성생물학 기술을 통해 대장균을 고수율 카로티노이드의 이형 생산을 위한 공학에 성공했다 [20-21].그러나 대장균이 파지에 감염되기 쉽고 [22] 내독소의 존재 등의 위험성 때문에 대장균을 이용해 리코펜을 생산하는 것은 현재 특정 식품 안전상의 위험을 안고 있어 산업적 응용이 제한적이다.
Saccharomycescerevisiae는 진핵생물 모델 생물로서 게놈 서열이 밝혀져 있고, 세포 생물학의 특징이 잘 밝혀져 있으며, 이를 위한 성숙한 유전자 조작 도구와 방법이 있다.Saccharomycescerevisiae의 대규모 발효시 파지 오염의 위험이 없으며, 일반적으로 대장균보다 안전한 것으로 판단된다.따라서 리코펜의 이형 생산을 위해 Saccharomyces cerevisiae를 변환하는 대사공학을 이용하면 응용 가능성이 클 것으로 판단된다.대장균처럼 Saccharomyces cerevisiae는 자체적으로 카로티노이드를 합성할 수 없고 관련 합성 유전자를 도입해야 한다 [23~26].
Yarrowia lipolytica는 대량의 지질을 생산하는 비전통적인 미생물 숙주이며 안전하다고 간주됩니다.카로티노이드를 직접 합성할 수는 없지만 전구체 아세틸 코엔자임 A를 다량 생성할 수 있으며, 외인성 핵심 효소를 도입하여 카로티노이드의 합성이 가능하다.연구자들은 MVA 경로를 통한 카로티노이드 생산을 위한 유망한 숙주로 간주되는 Lipomyces 효모를 공병하기 위한 많은 유전적 도구를 개발해왔다 [27-28].

진핵 미세조류는 독립영양미생물로서 빛에너지와 이산화탄소를 이용하여 바이오매스를 생산할 수 있으므로 테르페노이드의 지속가능한 생산을 위한 대사잠재력이 크다.하지만 현재 고준위 조류의 대사공학 연구는 다른 숙주에 비해 많이 뒤처져 있어 [29] 어느 정도 응용에 한계가 있다.
같은 붉은 이스트 Rhodosporidium toruloides 생산 할 수 있는 색소 β-carotene와 γ-carotene을 통해 세포 내 biosynthesis다.연구진은 배양 조건과 돌연변이 작용을 최적화해 카로티노이드 생산 능력을 향상시켰다.그러나 현재 적색 효모에 대한 연구는 매우 제한적인데, 이는 이용 가능한 유전체 데이터의 한계와 주요 유전자의 기능 주석 부족으로 인해 [30] 생산량이 많은 카로티노이드의 대사공학에 큰 걸림돌이 되고 있는 것으로 보인다.에탄올 축적 없이 고밀도로 생장할 수 있는 Pichia pastoris 등 카로티노이드를 생성하지 않는 효모들도 카로티노이드를 합성할 수 있도록 공학적으로 개발되었으나, 수확량이 적어 연구가 필요하다 [31].
3.2 미생물을 공학적으로 라이코펜을 합성하기 위한 전략
1) Upstream module (전구체 IPP/DMAPP 공급) 향상
리코펜과 같은 카로티노이드의 높은 수율을 얻기 위해서는 일반 전구물질인 IPP와 DMAPP의 합성을 높이는 것이 효과적인 전략이다.IPP와 DMAPP 합성에는 MEP 경로와 MVA 경로의 두 자연 경로가 포함된다.(a) MEP 경로는 주로 원핵생물에서 발견된다.DXS와 IDI는 일반적으로이 경로에서 주요 속도 제한 효소로 간주되며, [32] 아이소프레노이드 합성을 향상시키기 위해 과발현 되었다.Li 등 [33]은 IspA, ispH 및 ispE 가 DXS및 IDI 과발현 균주에서 경로의 플럭스를 더욱 증가시킨다는 것을 발견하였다.ispG의 과발현은 세포 내 MEC의 유출을 효과적으로 감소시켜 하류의 isoprenoid 생성을 크게 증가시킬 수 있다 [34].이를 바탕으로 Li 등 [35]은 IspG와 IspH를 활성화시켜 MEP 중간체의 축적을 제거함으로써 리코펜 생성을 77% 증가시키는 데 성공했다.(b) MVA 경로는 주로 진핵생물에서 발견된다.HMG-CoA 환원효소는 MVA 경로 (36)를 통한 이소 프레 노이드 화합물의 생합성의 첫 번째 단계이다.Saccharomyces cerevisiae에서 HMG-CoA 환원 촉매 영역 (tHMG1)을 과발현시키면 리코펜 (lycopene)의 생성을 증가시킬 수 있다 [24].또한, MEP 경로를 최적화하여 카로티노이드 생산에 어느 정도 진전이 있었지만, MEP 경로에서 자연 숙주의 조절 메커니즘은 그 적용에 한계가 있다 [37].이 경로를 우회하기 위해 주파인 등 [20]은 완전한 MVA 경로와 외인성 유전자를 대장균에 도입하였고, 회분식 공급 및 발효 최적화를 통해 1.44 g/L의 라이코펜 수율을 얻었다.
(2) downstream module (the lycopene heterologoussynthesis pathway)에 대한 연구.일반적인 전략은 heterologous 경로유전자를 카로티노이드가 아닌 숙주에 도입하여 카로티노이드를 생성하여 테르펜 합성 전구물질인 IPP와 DMAPP을 카로티노이드로 전환하는 것이다.Verwaal 등 [38]은 lycopene desaturase를 암호화하는 cDNA 뿐만 아니라 geranylgeranyl pyrophosphate synthase와 octahydro-lycopene synthase를 암호화하는 유전자를 포함하는 플라스미드를 대장균에 발현시켰으며, 궁극적으로 lycopene의 축적을 관찰하였다.CrtI 사본의 도입과 tHMG1 carotenoid-synthesizing 효모 세포에 β을 늘렸-carotene 콘 텐 츠 가 있습니다.이형경로 유전자의 높은 수준 및 유전적으로 안정적인 발현을 위해 Tyo 등 [39]은 화학적으로 유도된 염색체 진화를 위한 플라스미드 없는고 gene-copy 발현 시스템을 구축하였으며, 이를 engineered E. coli에 사용하였고 궁극적으로 플라스미드 발현 시스템에 비해 리코펜 생산을 60% 증가시켰다.연구에 따르면 리코펜 합성 경로를 최적화하는 것은 이형질 고수율 리코펜에 매우 중요한 것으로 나타났다.
3) 우회경로의 하향조절
4) 리코펜 합성의 전구체인 FPP는 많은 효모 대사산물 (유비퀴논, 테르펜알코올, 스쿠알렌 등)의 일반적인 전구체이기도하다.그러나 이러한 전구체 경쟁 경로 유전자 (예:스쿠알렌 합성 유전자)의 직접적인 녹취는 세포 성장에 큰 영향을 미칠 것이다.따라서, 많은 학자들은 리코펜의 합성 플럭스를 향상시키기 위해 이러한 경쟁 경로를 하향 조절하는데 전념하고 있습니다.자연에 대한 약 한 프로 모터를 대체 기획자를 downregulate 스쿠알렌 경쟁의 유전자 sqs1 titratable 생산량을 증가시 킬 수 있다고 synthase β-carotene에서 (453.9 ± 20. 2) mg/L을 (797.1 ± 한 이래) mg/L에 Yarrowia lipolytica [40]다.심원표 등 41명은 Saccharomyces cerevisiae에서 고포도당 유도/저포도당 억압 촉진제인 pHXT1 제제를 이용하여 배양액의 포도당 농도 변화에 따른 erg9 유전자와 카로티노이드 경로 유전자의 순차적 발현을 이루어내어 효모에서 리코펜 생성이 크게 증가하였다.홍 등 (42)은 Saccharomyces cerevisiae의 dpp1과 lpp1 유전자를 제거하여 farnesol의 생성을 위한 경쟁 경로를 억제하였고, lycopene 생성도 증가시킨 erg9발현을 하향조절하여 ergosterol 생성을 억제하였다.위의 연구는 경쟁 경로를 하향 조절하는 것이 리코펜 생산을 늘리는 데 효과적인 전략임을 충분히 증명했다.
4) 섀시 셀의 변환
리코펜 이형 합성 경로를 최적화하는 것 외에도 숙주 섀시 세포의 형질전환도 이형 경로와 일치하도록 요구됩니다.섀시세포의 개량은 아세틸-coa 전구체의 플flux 강화 [43], ATP와 NADPH와 같은 보조인자 공급 강화, 특정 비필수 유전자 때려잡기, 변종의 적응적 진화 등이 있다.아세틸-coa는 카로티노이드 생합성을 위한 기질이다.천옌 등은 Saccharomyces cerevisiae에서 ypl062w 유전자의 작용 메커니즘을 상세히 연구하였다.ypl062w의 결실은 아세틸 코엔자임 A의 플럭스를 향상시키고 궁극적으로 최대 1.65 g/L인 리코펜의 생성을 증가시킬 수 있습니다.Zhou 등 (26)은 대사공학 기술과 결합된 Saccharomyces cerevisiae의 적응적 진화를 이용하여 8.15 g/L lycopene의 회분식 발효 수율을 달성하였다.에너지로서의 ATP와 전력을 감소시키는 NADPH의 공급은 카로티노이드 합성에 영향을 미치는 중요한 요소이다.중앙 대사 모 듈을 수정 하여 탄소 원본에 대한 동화 (EMP과 PPP 경로), ATP와 NADPH의 공급은 향상 된이었고, 조작 된 대장균을 합성 할 수 있는 2. 1 g/L β-carotene에 배치 발효 [44].sucAB과 sdhABCD 유전자의 발현을 조절하면 TCA 회로의 탄소 플럭스를 증가시키고 ATP의 공급을 증가시킬 수 있다.또한 talB 유전자를 조절하면 NADPH의 공급을 증가시킬 수 있으며, 이는 대장균에 의한 리코펜 합성 수율을 3.52 g/L로 증가시킨다 [45].
(5) 고수율의 리코펜을 생산하기 위한 Saccharomyces cerevisiae의 체계적인 대사공학.요약은 그림 2와 3에 나와 있습니다.
시빈 등 (25)은 대사공학을 통해 리코펜을 효율적으로 생합성할 수 있도록 사카로마이세스 세르비지애 (Saccharomyces cerevisiae)를 체계적으로 공학하였으며, 4가지 주요 문제를 제시하였다:① 2차 대사산물의 축적과 숙주세포 성장이 균형을 이루도록 해야 한다;(b) 효모에서 리코펜의 이형 합성 경로를 향상시킬 필요가 있다;(c) 효모 섀시 셀은 더 많은 전구체 물질 및 감소 전력을 제공하기 위해 변형되어야;(d) 효모 발효 기술을 최적화해야합니다.그리고, (a) 라이코펜 이형 합성 경로를 합리적으로 제어할 수 있는 GAL 프로모터 그룹을 선별하여 시간 서열면에서 효모 세포 성장을 라이코펜 생성물 축적으로부터 분리하고, 프로모터 강도 또한 생성물 합성 기간 동안 구성적인 강한 프로모터와 비교할 수 있는;(b) 라이코펜 이형 합성 경로의 세 가지 주요 유전자원을 종합적으로 스크리닝 하여 Saccharomyces cerevisiae에서 효율적으로 기능하는 PaCrtE, PagCrtB 및 BtCrtI의 새로운 최적 조합을 얻었다.(c) 또한 Saccharomyces cerevisiae섀시 세포에서 리코펜 합성에 필요한 충분한 전구물질, 아세틸계 코엔자임 A 및 환원력 NADPH (reduced co효소II)를 제공하고, 리코펜 축적에 영향을 미치는 특정 내인성 비필수 유전자를 제거하여 리코펜 생산을 더욱 증가시키기 위해 일련의 개량을 수행하였다;(d) 이러한 시스템 대사공학 방법을 통해 Saccharomyces cerevisiae발효 공정의 변환과 함께 리코펜을 효율적으로 생합성한다.
그는 △ 2차 대사산물의 축적과 숙주세포 성장이 균형을 이루어야 한다;(b) 효모에서 리코펜 이형 합성 경로를 향상시킬 필요가 있습니다;(c) 효모 섀시 셀은 더 많은 전구체 물질 및 감소 전력을 제공하기 위해 변형되어야;(d) 효모 발효 기술을 최적화해야합니다.그리고, (a) 라이코펜 이형 합성 경로를 합리적으로 제어할 수 있는 GAL 프로모터 그룹을 선별하여 시간 서열면에서 효모 세포 성장을 라이코펜 생성물 축적으로부터 분리하고, 프로모터 강도 또한 생성물 합성 기간 동안 구성적인 강한 프로모터와 비교할 수 있는;(b) 라이코펜 이형 합성 경로의 세 가지 주요 유전자원을 종합적으로 스크리닝 하여 Saccharomyces cerevisiae에서 효율적으로 기능하는 PaCrtE, PagCrtB 및 BtCrtI의 새로운 최적 조합을 얻었다.(c) 또한 아세틸코엔자임 A 및 환원력 NADPH (환원된 코엔자임 II)와 같은 리코펜 합성에 필요한 충분한 전구체를 제공하기 위해 Saccharomyces cerevisiae섀시 세포에 일련의 개량을 수행하였으며, 리코펜 축적에 영향을 미치는 특정 내인성 비필수 유전자를 제거하여 리코펜 생산을 더욱 증가시켰다;(d) 이러한 시스템 대사공학적 방법을 통하여 Saccharomyces cerevisiae에 의한 합성매체 발효의 최적화와 함께 리코펜 수율이 3.28 g/L로 높게 나타나 초기에 산업적 수준에 도달하였다.의 규모를 현재, Chinese-style 발효 6, 000까지 성공적으로 조정 되었습니다. l. 게다가, 신진대사 공학의 전략을 위해 개발로 미세는 또한 성공적으로 확장 되었습니다 biotechnological 합성의 다른 terpene 화합물, 같은 β-farnesene [46], bergapten [47], 그리고 같은 다른 sesquiterpenes β-caryophyllene다.그 중 등샤오민 등 [47]은 이러한 시스템 공학적 전략을 이용하여 Saccharomyces cerevisiae 균주의 대사공학으로 생산된 bergapten의 생산량을 34.6 g/L로 증가시켰다.
리코펜 조작 세균의 발효과정 3.3
현재 미생물에 의한 고수율 라이코펜의 발효에 대한 연구도 큰 진전을 보이고 있다.균주마다 사용하는 발효 원료, 공정 및 척도는 다를 것이다 (표 1에 요약). 미생물의 종류에 따라 발효 공정에 사용되는 기질 기질은 다르다.보다 성숙한 리코펜 이형합성을 한 발효 숙주는 일반적으로 Escherichia coli와 Saccharomyces cerevisiae이다.일반적으로 대장균 발효물은 글리세롤을 탄소원으로 사용하며 [20, 45], Saccharomyces cerevisiae는 포도당을 주로 탄소원으로 사용한다 [23~26].
주패인 등 [20]은 가공된 대장균을 위한 탄소원으로 글리세롤을 사용하였고 5 L 발효기에서 완전 합성 매질을 이용하여 1.44 g/L의 리코펜 수율을 얻었다.이후 발효배율은 150 L로 확대되어 1.32 g/L를 산출하여 균주의 생산을 위한 배지 확장이 가능함을 알 수 있었다.Sun 등 [45]은 대장균을 회분식 공급에서 글리세롤을 탄소원으로 하는 7 L 발효기에서 발효하도록 조작하여 최종적으로 3.52 g/L의 리코펜을 얻었다.천옌 등 (24)은 제조된 Saccharomyces cerevisiae를 이용하여 포도당과 에탄올을 탄소원으로, 효모 추출물과 펩톤의 질소원을 회분식 공급법을 이용하여 5 L 발효기에서 발효시켰다.

1.65 g/L의 리코펜 역가를 얻었다.시빈 등 25)은 제조된 Saccharomyces cerevisiae를 이용하여 포도당과 에탄올을 탄소원으로, 황산암모늄을 질소원으로 사용하는 7 L 발효기에서 2단계 fed-batch 발효를 수행하였다.발효 국물 중의 포도당과 에탄올 잔량을 엄격하게 관리하여 최종 3.28 g/L의 리코펜 역가를 얻었다.Saccharomyces cerevisiae는 대장균에 비해 높은 식품 안전성 및 파지 감염에 대한 저항성 등 많은 장점을 가지고 있으므로, Saccharomyces cerevisiae에 의한 리코펜의 발효에 대한 연구가 더욱 유망하다.현재는 펩톤과 효모추출물을 효모 발효를 위한 질소원으로 자주 사용하는 천연 YPD 매체 [24, 26]와 황산암모늄, 효모추출물, 펩톤을 혼합 질소원으로 하는 반합성 매체 [23], 황산암모늄을 질소원으로 하는 완전 합성 매체 [25] 등이 있다.완전 합성 매체는 저렴하고, 반복 발효와 스케일업이 용이하며, 추후 최적화에 편리한 명확한 구성을 갖는 장점이 있다.앞으로 리코펜 생산을 높고 안정적으로, 반복 가능하고, 확장 가능하게 하기 위한 합성 효모 매체의 발효에 대해 더 많은 연구가 이루어져 후속 산업 응용을 위한 견고한 토대를 마련해야 합니다.
미생물에 의해 합성한 리코펜의 추출 및 정량 3.4
다음과 같은 카로티노이드리코펜은 강한 항산화 특성을 가지고 있습니다, 그리고 추출 과정에서 산화와 이성질화의 위험을 최소화해야 한다 [49].예를 들어, 일부 연구에서는 빛이 보호되는 조건 [20, 45]에서 작동하거나, 추출제에 항산화제 BHT를 첨가하는 방법을 선택했습니다 [24-25, 27].일반적으로 사용되는 추출 용매로는 아세톤, 석유 에테르, 클로로포름, 헥산, 에틸 아세테이트 등이 있다 [49].리코펜은 세포 내 생성물이기 때문에 세포벽 파괴가 필요하며, 숙주 세포벽의 두께에 따라 세포벽 파괴 방법이 달라진다.예를 들어, 대장균의 세포벽은 비교적 얇아 일반적으로 아세톤 소생을 사용하고, 이후 55 °C의 수조에서 세포벽 파괴 (20, 45;지용성 효모 (lipid-soluble 효모)나 사카로마이세스 세르비지애 (Saccharomyces cerevisiae)와 같은 진핵생물은 세포벽이 더 두껍고, 유리구슬과 추출시약은 보통 흔들어서 세포를 부수기 위해 첨가된다 (25, 27;염산을 넣은 수조에서 끓이면 효모 세포벽이 깨지기도 한다 [23~24].
리코펜과 같은 카로티노이드를 검출하고 정량하기 위해 사용되는 방법도 기존 연구에서 다양하다.대부분의 연구 고성능 액체 크로마토그래피 (HPLC)를 사용 하여 리코 펜 같은 카로 티 노이 드을 감지하고 β-carotene [23-26, 45]., 사용하는 동안 몇 ultraviolet-spectrophotometry [20, 27].왜냐하면 UV-spectrophotometer 검출은 동일한 흡수파장을 가진 불순물에 의해 간섭을 받고, HPLC 방식은 먼저 다른 물질을 분리한 후 흡수값을 검출하여 검출하기 때문입니다.상대적으로 HPLC에 의한 리코펜과 같은 카로티노이드의 정확한 정량이 더 정확하며, UV-spectrophotometer 검출은 발효 중 수율 변화 추이를 초기에 평가할 수 있는 보조 수단으로 사용될 수 있다.

대부분의 연구 표준 곡선을 사용 해당 리코 펜과 β-carotene 수율을 계산하는 기준 [23-26, 45]., 하지만 리코 펜을 구입 했 는지 여부를 나타내지 않/β-carotene 기준을 보정 했다.표준곡선을 그리기 전에 [25] 표준용액의 농도를 보정했다고 명시적으로 밝힌 연구자는 극히 일부에 불과했다.이유는 준비 된 표준의 용액의 농도를 보정 해야하는 것은 그것이 어렵 다는 것을 정확하게 무게 가 소량의 리코 펜 또는 β-carotene 표준, 그리고하 는지 여부를 확인하는 것은 쉽지 않 정확하게 리코 펜 결정체 유기 용매에 용해 완전히 가지고 있다.또한, 보관 방법 및 시간에 따라 구매한 표준의 순도도 변경될 수 있습니다.이러한 객관적 인자들은 리코펜 표준곡선의 그리기에 큰 오차를 발생시켜 계산된 수율이 매우 정확하지 않을 수 있다.상기 인자들의 간섭을 제거하기 위해서 먼저 분광광도계를 이용하여 제조된 리코펜과 다른 카로티노이드 표준용액의 흡광도를 측정한 후, 해당 소멸계수 (25, 50-51)에 따라 용액 중의 카로티노이드의 절대함량을 계산하는 것이 일반적인 방법이다.이 방법은 무게가 부정확하거나 시료의 불완전한 용해로 인한 오류를 제거할 수 있습니다.요약하면 HPLC를 이용하여 시료중의 리코펜을 정확하게 검출하고 보정된 표준용액을 이용하여 표준곡선을 그려보면 정량결과가 더욱 정확할 것이다.
4 결론 및 전망
리코펜은 강한 항산화제로서 여러가지 좋은 생리기능을 갖고있어 시장전망이 넓다.본 논문은 라이코펜의 물리화학적 특성, 생리학적 기능 및 생산방법에 대한 상세한 고찰과 미생물 숙주세포의 다양성 선발, 최신 대사공학적 전략, 발효방법 및 라이코펜 추출 및 정확한 정량방법 등 생명공학에 의한 라이코펜 생산에 대한 현재 연구 진행상황을 요약하는데 중점을 두고 있다.
비록 라이코펜의 생명공학적인 합성에 어느 정도 진전이 있었지만, 라이코펜 생합성 과정은 많은 영향인자가 있는 복잡한 공학 연구 프로젝트이기 때문에 여전히 많은 문제점이 있다.다음은 향후 관련 연구 방향을 요약하면:
(1) 발효 생산의 규모화 및 안정적인 복제.현재 라이코펜의 생명공학적 합성에 대한 대부분의 연구는 아직 소규모 발효탱크 실험의 실험실 단계에 머물러 있으며, 산업 연구는 톤 또는 수십 톤 단위로 계산된 대규모 발효 생산에 기반하는 경우가 많다.소규모에서 시험생산으로의 발효규모화는 단순히 선형적인 탱크부피의 증가가 아니라 균주의 성장모델의 변화와 함께 불균일한 열전달, 물질전달, 산소전달 등의 많은 과제를 수반한다.작가 &에 따르면#39;s 실제 경험상, 조작된 균주의 발효 증폭 과정 중, 조기 균주 노화, 균주 표현형 저하, 공급 전략의 변화, 불안정한 발효 수율 등의 문제가 발생할 수 있다.연구자들은 발효 공정 증폭의 매개 변수와 조건을 개별적으로 조정할 필요가 있습니다.향후 발효생산 규모의 증폭에 대한 연구는 생명공학을 이용한 리코펜의 공업적 생산 문제를 해결하는 관건이다.
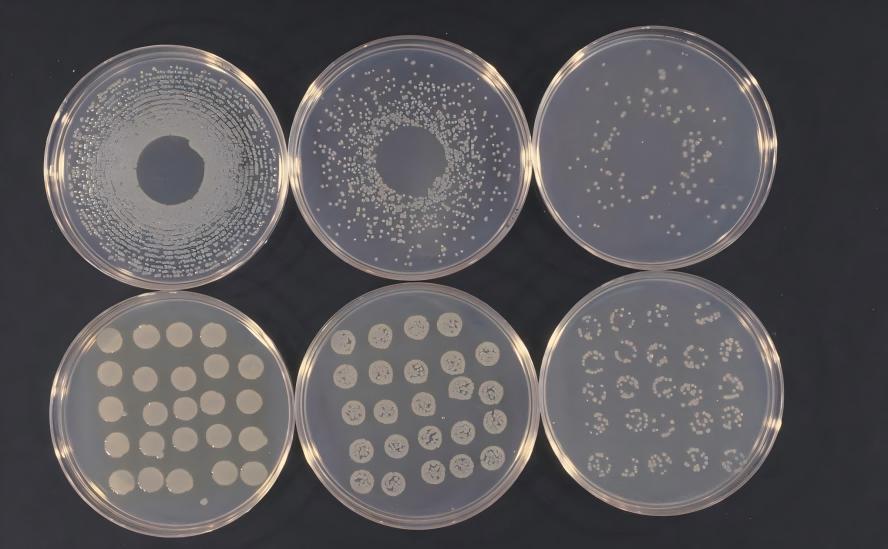
(2) 미생물원으로부터 리코펜의 추출 및 정제과정.리코펜은 세포 내 생성물로 추출 및 정제 과정은 세포 파괴, 불순물 제거, 리코펜 결정화 등의 많은 단계를 거칩니다.이 과정에서 리코펜은 쉽게 산화되고 이는 구조적인 변화를 가져온다.따라서 추출율을 보장하기 어려우며, 미생물 원료로부터 리코펜의 추출 및 정제과정에 대한 심층적인 연구가 필요하다.
(3) Research on quality testing of 미생물이lycopene. Although microbial lycopene is also a product of enzymatic catalysis, it is not derived from natural tomatoes and may involve issues such as genetic modification. Therefore, microbial lycopene must first be structurally identified to ensure that it is consistent with natural tomato sources, and then quality testing of product quality such as heavy metal residues and microbial content is required. Ensuring the structure and quality of the product is also an important factor affecting the market application of microbial lycopene.
(4) Production cost control: If it is to compete in the market with naturally extracted lycopene, biotechnologically synthesized lycopene must have a significant cost advantage. The main costs of biologically fermenting to produce lycopene include 발효raw materials, depreciation of equipment, extraction and purification, labor, and marketing. Cost factors must be considered when designing and optimizing production 프로세스conditions, such as using cheaper fully synthetic fermentation media, spreading costs by scaling up fermentation, and using more advanced methods such as enzymatic cell disruption to extract lycopene to reduce production costs.
이러한 문제를 해결하는 것은 생명공학에 의한 리코펜 생산의 산업화를 촉진하는데 이론적, 실무적으로 매우 중요하며, 또한 다른 고부가가치 천연물의 생명공학 생산에 대한 연구의 참고자료를 제공할 수 있다.
참조
[1]에 대한 MK-4의 보호 효과 유리 디스 >의 산화를 ⁃ enoids 위액 모의 [J]에 있다.저널의 Huazhong 발견 ⁃ 문화 대학, 2020년 39 (2):102-111 ( 중국 with English abstract)를 참고하십시오.
[2] 류 KY, 류 X Y, 왕 Q, et 알다. 최적화 의 주요 재료 비율 of 화합물 과일 and 야채 d-최적 혼합물 설계에 의해 리코펜이 풍부한 와인 [J.중국 양조 ⁃ ing, 2022년 41 (2):164-169
[3] ZARDINI A A,MOHEBBI M,FARHOOSH R 외 생산 and 성격 묘사 of nanostructured 지질 리코 펜이들어 있는 항공사와 고체 지질 나노 입자을 음식 fortifi ⁃ cation다 [J다] 저널 음식의 과학 and 기술,2018,55 (1):287-298.
[4] LEE MT,CHEN B H. 가열 중 리코펜의 안정성 그리고 모형 시스템 [J]에서 조명.식품화학,2002, 78(4):425-432.
[5] 왕 X W, 시아이 B, 왕 Kq. 천연 리코펜 [J]의 안정성.Hunan Agricultural University 논문집 2002,28(1):57-60 (in Chinese with English abstract).
[6]MA T G.Physiological functions and application 리코 펜 [J]의다.시리 얼 &2008년 기름, 21일 (1):46-48 (Chi ⁃ nese with English abstract)를 사용한다.
LI J,YAN W,LIU Y H 등 리코펜의 건강기능 및 응용에 관한 연구 진행.농업과 기술,2016,36(15):5-6 (중국).
[8]JIANG L H,LIU H 의 항산화능력에 대한 연구 진행 F,HAO G F,et astaxanthin다 [J다]식품산업과학기술,2019, 40(10):350-354.
[9]PENG Y J, LU H P, WANG S N, 외 자연물에 대한 연구 및 전망을 제시한다 astaxanthin다 [J다]중국 음식 첨가물, 2017년 (4):193-197.
[10] ASSAR E A, VIDALLE M C, 초 프라 M, et al.Lycopene 행위의 억제를 통해나는 κ B, B 산화효소 NF 하지 않 으려면-κ 신호 ⁃ ing에서 인간의 전립선과 유방암 세포 [J]이다.종양 biolo ⁃ gy, 2016년, 37 (7):9375-9385.
[11]는 라오 A V,은' s. 역할 항 산화의 lycopene 암과 심장병에서 [J.미국영양학회지,2000,19(5):563-569.
[12]HUO S X,YANG Q S,YUE X H, 기타. 리코펜 함량 측정방법 토마토 껍질 [J.외식산업,2019,40(6):263-265 (영문 초록 포함).
[13] 카로티노이드 및 새로운 중간체의 제조를 위한 KARL M.Method:US5208381[P].1993-05-04.
[14] KIRBY J,KEASLING JD.Biosynthesis of plant isoprenoids:perspectives. 「 식물이소프레노이드의 생합성 」 (영어) 을 microbial 공학 [J]다.연간 검토 of 식물생물학 (plant biology),2009,60:335-355.
[15] ROHMER M KNANI, M, SIMONIN P, et al.Isoprenoid 바이오 ⁃ 합성 박테리아:에서 소설 경로를 일찍 계단 ⁃ ing to isopentenyl diphosphate다 [J다]이 생화학 저널, 1993,295(Pt 2):517-524.
[16] 블로흐 K, 차이킨 S, 필립스 A H, 그 외 메발론산 피로인산염, 이소펜테닐피로인산염 [J].Journal of biological chemistry,1959,234(10):2595-2604.
[17] CHOUDHARI S M, ANANTHANARAYAN L, 노래, ⁃ HAL R s. en ⁃을 위한 대사 stimulators과 억제 제의 사용 hanced β의 생산에 의해-carotene 및 리코 펜 Blakeslea 트라이 ⁃ 스포라 NRRL 2895와 2896[J].바이오원천기술,2008, 99(8):3166-3173.
[18]ANATES T M R,DE CASTROTES A E, PEREZTES J c. 개선. method of producing 리코 펜, 준비 을 얻기 리코 펜, 그리고 application 다:CN1617934A [P].2005-05-18다.
[19] 왕 J F, 류 X J, 류 R S의 et al.Optimization, 매트 ⁃ ed fermentation process 을 the production of lycopene by Blakeslea trispora NRRL 2895(+)와 NRRL 2896 (−) [J]다.Bioprocess and biosystems 2012년 공학, 35 (4):553-564.
[20] ZHU F Y,LU L,FU S,et al.Targeted 공학과scale up of lycopene 과잉 생산in Escherichia coli (J.공정생화학,2015,50(3):341-346.
[21] 공원 S Y BINKLEY R, M, 김 W J, et al.Metabolic에 기술부 ⁃ neering 대장균의 대한 고위급 astaxanthin produc ⁃ 생산성이 높은 tion [J.대사 공학,2018, 49:105-115.
[22] RAY B L,RAETZ C R. 4&를 통합하는 Escherichia coli 막에서 gram-negative endotoxin.A novel kinase의 생합성#39;-지질 A[J]의 인산염.1987년 Journal of biologi ⁃ 칼 화학, 262 (3):1122-1128다.
[23] 장 W P LV X, M, 여러분은 L D, et al. 시공 리코 펜-overproducing로 미세 결합 함 으로써 직접적인 진화와 대사 ⁃ 에드 공학 [J]다.2015년 대사 엔지니어 ⁃ ing, 30:69-78다.
첸 [24] Y, 샤 오 W H 왕 Y, et al.Lycopene overproduc ⁃ 졌을 통해로 미세에 경로 공학 (경로engineering)과 호스트 공학 (host engineering)을 결합 [J/OL].사실상 ⁃ 미생 물의 세포 ries, 2016년, 113 15 (1):[2023-03-13]다.토리당 https://microbialcellfac ⁃다. biomedcentral다. com/articles/10.1186/s12934-016-0509-4다.
[25] SHI B,MA T,YE Z L 등 체계적인 대사공학 Saccharomyces의 cerevisiae for lycopene overproduction [J]다.한국농식품화학학회지 2019년,67(40):11148-11157.
조우 [26] K, 유 C, 량 N, et 적응 알다. 진화 그리고 대사 공학적 리코 펜 생산 증대에 Saccharo ⁃ myces cerevisiae 향상 된 선임자들을 통해 공급과 활용 [J]이다.한국농식품화학학회지,2023,71(8):3821-3831.
[27] 가오 S L, 통이 Y, 주 L 등의 반복 통합 multiple-copy 경로 유전자 Yarrowia lipolytica을 heter ⁃ ologous β-carotene 생산 [J]이다.대사 공학, 2017,41:192-201.
[28] LARROUDE M,CELINSKA E,BACK A,et al. 변형시키기 위한 합성 생물학 접근 com로 Yarrowia lipolytica ⁃ petitive biotechnological 프로듀서의 β-carotene [J]다.115 Biotechnol ⁃ ogy 그리고 생명공학, 2018년, (2):464-472다.
[29] GUERIN M 헌 틀리, M E, OLAIZOLA M.Haematococ ⁃ cus astaxanthin:응용 프로그램을 인간의 건강과 영양 [J]다.생명공학동향,2003,21(5):210-216.
[30] 뮤 디아스 C, 실바 C, FREITAS C, et al.Effect의 중간에 pH Rhodosporidium toruloides NCYC 노 921 드 및 립 ⁃ id production 평가 by 흐름 cytometry다 [J다]적용 바이오 ⁃ 화학 그리고 생명공학, 2016년, 179 (5):776-787.
[31] BHATAYA A, SCHMIDT-DANNERT C,리 P. C. 메타 ⁃ bolic 공학의 Pichia pastoris-33 리코 펜을 produc ⁃ 졌다 [J다]공정생화학,2009,44(10):1095-1102.
[32] YANG J M,GUO L Z. 생합성 의 β-carotene에 엔지니어링 부서 ⁃ neered 대장균 유럽의회 의원과 영국 측 길 [J]를 사용하고 있다.2014년 Microbi ⁃ 알 셀 공장, 13:160다.
[33]리 Y F,린 Z Q, 황 C, et al.Metabolic 공학을 이용한 대장균 CRISPR-Cas9 명상 [J] ⁃ 편집 게놈을 가지고 있다.대사공학 2015년, 대사공학 31:13-21.
[34] 저 우 K, 추읍 R Y, STEPHANOPOULOS G, et al.Metab ⁃ olite 프로 파일 링 methylerythritol 확인 cyclodiphosphate ef ⁃ 플럭스를 제한하는 단계 로서 미생물 isoprenoid 생산 [J]이다.플로스원,2012,7(11):e47513.
[35]리 Q Y, 팬 F Y 가 오 X, et IspG의 al.Balanced 등록과 IspH 유럽의회 의원을 제거 하기 위한 중간 및 메신저 축적 ⁃ 증명 isoprenoids 생산에서 대장균다 [J다]대사공학 2017년, 대사공학 44:13-21.
[36] DIMSTER-DENK D, THORSNESS M K, RINE J. 먹이의 규제 가 다시 ⁃ 3-hydroxy-3-methylglutaryl 코엔 자임 한 회신 ⁃ ductase로 미세 [J]에 있다.Molecular 생물학of the cell,1994,5(6):655-665.
[37] 마틴 V J J, PITERA D J, 시들 S T, et al.에 기술부 ⁃ neering mevalonate 경로인 대장균에 produc ⁃ tion of terpenoids다 [J다]자연 생명공학, 2003년, 21일 (7):796-802다.
[38] VERWAAL R,WANG J,MEIJNEN J P,et al. 고수준 생산 of beta-carotene in Saccharomyces cerevisiae 의에 의해 연속적인 혁신 carotenogenic 유전자를 잰은 ⁃ thophyllomyces dendrorhous다 [J다]적용 and 환경미생물학,2007,73(13):4342-4350.
[39] TYO K E J,AJIKUMAR P K,STEPHANOPOULOS g. 안정 유전자 중복 가능하게 장기적인 selection-freeheterologous pathway [J] 표현이다.자연 생명공학, 2009,27(8):760-765.
[40] KILDEGAARD K R, ADIEGO-PEREZ B,도 메 네 크 BELDA D, et al. 공학의 Yarrowia lipolytica을 프로 ⁃ duction의 astaxanthin다 [J다]합성 및 시스템 biotechnolo ⁃ gy, 2017년, 2 (4):287-294.
[41] 장 W P, 여러분은 L D, 루 X M, et al.Sequential 생합성 균형 잡 힌의 활용을 위한 대사인 터 밀 란의 통제 ⁃ 중재로 미세 [J]에 있다.2015년 대사 엔지니어 ⁃ ing, 28:8-18다.
[42] HONG J,PARK S H,KIM S,et al.Efficient production of lycopene in Saccharomyces cerevisiae by enzyme engineering and 막 증가 유연성을 그리고 NAPDH 생산 [J]다.한국미생물학회지 Applied microbiology and biotechnology,2019,103(1):211-223.
[43]리 J R,린 J Y, Z Y리, et al.Mining 향상 시키는 것을 규제하는 아세 트 산 stress-responsivegenes 리코 펜 syn ⁃ 논문:재조합 사카로마이세스 세레비지애 (Saccharomyces cerevisiae) [J/OL.마이크로 ⁃ biology 중국, 2023:1-24다 [2023-03-13다]
[44] 자 오 J, LI Q Y, 태양 T, et al.Engineering 중앙의 신진대사 모 듈 대장균 향상을 위한 β-carotene produc ⁃ 졌다 [J다]대사공학,2013,17:42-50.
[45] SUN T,MIAO L T,LI Q Y, 등 대사 공학에 의한 리코펜 생산 Escherichia 콜리다 [J다]Biotechnology letters,2014,36(7):1515-1522.
[46] 예 Z L,시 B, 황 Y L, 외. 미생물 발효 파르네센에서 시작하여 이소균 [J/OL]까지 비타민 E 생성의 혁명.The innovation,2022,3(3):100228[2023-03-13].https://doi.org/10.1016/j.xinn.2022.100228.
[47] 등 X M,시 B, 예 Z L, 등의 systematic identification of Ocimum sanctum sesquiterpenoid synthases and (−)-eremophilene overproduction in engineered yeast[J].신진대사에 기술부 ⁃ neering, 2022년, 69:122-133.
[48] 루오 Z S 류 N,'는 Z, et al.Enhancing isoprenoid syn ⁃ 논문에서 Yarrowia lipolytica isopentenol 사용 률을 표현 함 으로써 pathway and 바 꾼 세포 내 hydrophobicity다 [J다]대사공학,2020,61:344-351.
[49] CHOKSI P M,JOSHI V Y.A review on lycopene:추출, 정화, 안정성 및 응용 [J].국제 저널 식품특성 중,2007,10(2):289-298.
[50] 비안 G K, 마 T, 류 T G. in vivo 플랫폼으로 테르페노이드 과잉 생산 and the 세대 of chemical 다양 성이 [J]다.효소의 방법 (Methods in enzymology),2018,608:97-129.
[51] 스 콧 K j. 탐지 and 측정 of 카로 티 노이 드 UV/VIS spectrophotometry에 의해 [J.현재 음식에서 프로토콜 아나 ⁃ lytical 화학, 2001년, F2 키. 2 (1):-10이다.




